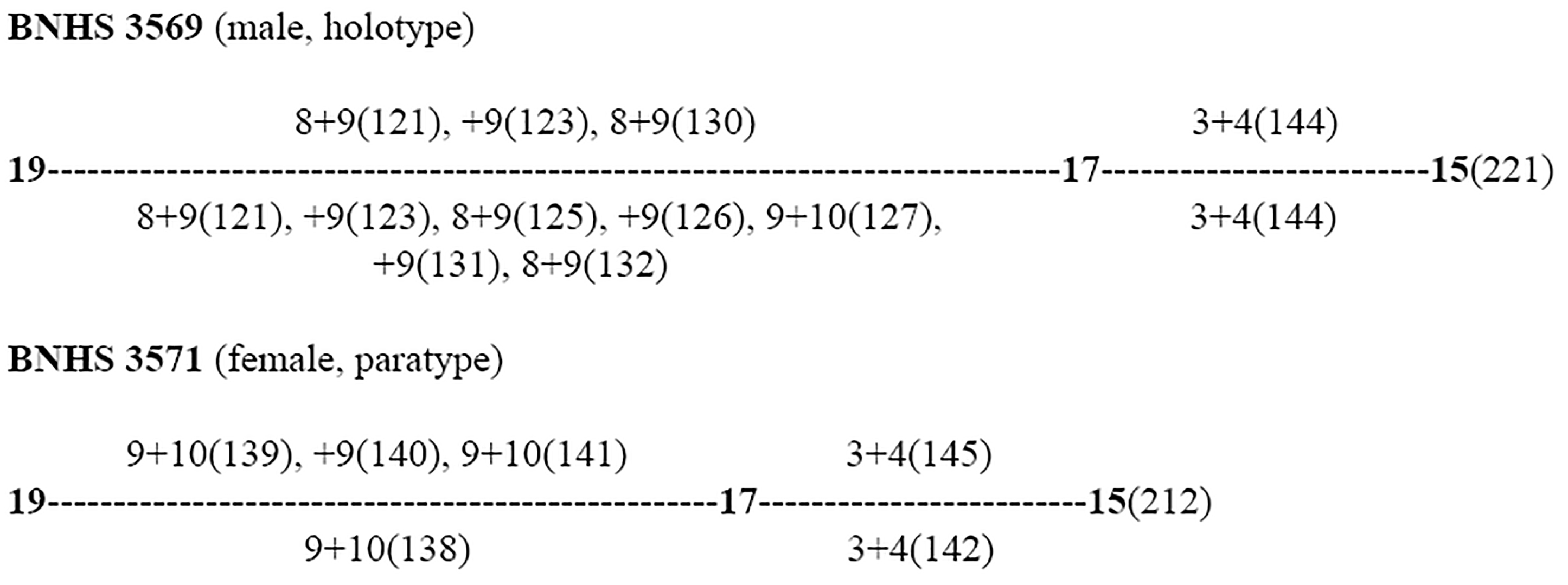Research Article |
|
Corresponding author: V. Deepak ( veerappandeepak@gmail.com ) Academic editor: Uwe Fritz
© 2023 Surya Narayanan, Sandeep Das, Y. Muhammed Anvar, Frank Tillack, Pratyush P. Mohapatra, David J. Gower, K. P. Rajkumar, V. Deepak.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Narayanan S, Das S, Anvar YM, Tillack F, Mohapatra PP, Gower DJ, Rajkumar KP, Deepak V (2023) On the taxonomic validity of Boiga whitakeri Ganesh et al., 2021 with new insights on Boiga dightoni (Boulenger, 1894) (Reptilia: Squamata: Colubridae). Vertebrate Zoology 73: 1-21. https://doi.org/10.3897/vz.73.e97002
|
Abstract
Colour polymorphism has been previously reported in several colubrid snakes including Boiga spp. In this paper, we report colour variations within the poorly known southern Indian Boiga dightoni, provide the first molecular data for this species, from two localities (including the type locality) and compare them with data from other congeners. Additionally, we provide detailed dentition and hemipenis descriptions for B. dightoni. Molecular data for B. dightoni show very little difference (0.2–0.4% 16S; 0.9–1.2% cyt b) to the recently described Boiga whitakeri, also from southern India. We have re-examined and present new information on the pholidosis of the type specimens of B. whitakeri and reconsider its taxonomic status. On the basis of molecular data and overlapping morphological characteristics, we argue that Boiga whitakeri and Boiga dightoni are conspecific, and place B. whitakeri under the subjective synonymy of the latter. Furthermore, we show that colour polymorphism in B. dightoni is a gender-independent character and that both colour morphs are found in high as well as low elevations and partly in sympatry. A revised key to the Boiga ceylonensis complex is provided.
Keywords
Boiga ceylonensis complex, taxonomy, synonymy, Kerala, Tamil Nadu, India
Introduction
The colubrid snake genus Boiga Fitzinger, 1826 is represented by 37 currently recognised species distributed from the southern Palaearctic and the Oriental region to the northern and eastern coasts of Australasia (
During our recent fieldwork in the southern Western Ghats, we collected two individuals of Boiga sp., one from Peermed and the other from Arippa, Kerala. The specimen from Arippa superficially resembled the holotype of B. whitakeri, in colour and inconspicuous dorsal markings, and the specimen from Peermed resembled the paratype of B. whitakeri in having prominent dorsal bands. We generated molecular and further morphological data for these two individuals and compared them with the types and with non-types of other Boiga spp. from the Western Ghats. In this work, we reassess the taxonomic status of B. whitakeri in light of new data on scale variation, and we report colour polymorphism within Boiga dightoni.
Materials and Methods
Molecular phylogenetics
We generated DNA sequences for two Boiga sp., a specimen (ZSI-CZRC-V-7541) from Peermed, Kerala (9.602710°N, 76.937857°E, 1238 m Above Sea Level (ASL)) approximately 9 km from the type locality of B. dightoni and one more specimen (BNHS 3617) from south of Shencottah gap (Arippa, Kerala, 8.831640°N, 77.038542°E, 195 m ASL) (Fig.

We extracted genomic DNA from liver samples stored in absolute ethanol at –20°C, using the DNeasy (QiagenTM) blood and tissue kit following the manufacturer’s protocol. We amplified partial sequences of two mitochondrial genes, 16S rRNA (16S) and cytochrome b (cyt b). Respective primers for these genes are as follows: 16Sar-L and 16Sbr-H (
Bidirectional sequences were checked manually using CHROMAS (http://technelysium.com.au/wp/chromas) and aligned using ClustalW with default prior settings implemented in MEGA 7 (
Maximum Likelihood (ML) analysis was performed using IQ-TREE (
Morphology
We examined 33 specimens of Boiga spp., including Boiga dightoni (n = 9), B. whitakeri (n = 2), B. nuchalis (n = 10), B. thackerayi (n = 4), B. flaviviridis (n = 1) and B. ceylonensis (n = 7) (Appendix
The numbers of dorsal scale rows are reported for one head length behind the head, at midbody (i.e., at the level of the ventral plate corresponding to half of the total ventral number), and at one head length anterior to the vent respectively. Dorsal scale row reduction formulae were based on
The following measurements were taken: snout-vent length (SVL); tail length (TL); head length (HL: distance between posterior edge of last supralabial and tip of the snout); head width (HW: at angle of jaws); head depth (HD: height at the occipital region); Frontal length (FL: at the longest point); frontal width (FW: at the widest point on the anterior region); eye diameter (ED: horizontal diameter); eye to nostril distance (E–N: anterior corner of eye to posterior edge of nostril); eye to snout distance (E-S: anterior corner of eye to tip of snout); frontal to snout (FrSN: anterior end of frontal to tip of snout); inter-orbital distance (IO: measured at the anterior edge of eyes); number of dorsal scale rows (DSR). All linear measurements, except SVL and TL were taken using Mitutoyo dial vernier callipers (to 0.1 mm). SVL and TL were measured using a thread and metal scale (to 1 mm).
Among the specimens checked in this study, the holotype (BNHS 3597) of Boiga whitakeri is in a poor state of preservation and the paratype (BNHS 1863) of this species is also damaged, especially its anterior ventral scales. Thus, the number of ventral scales provided here for the paratype of Boiga whitakeri (BNHS 1863) is not complete Appendix
Location records for both Boiga dightoni and B. nuchalis used for the map (Fig.
To obtain counts of teeth by a non-invasive procedure, the head of the holotype of Boiga dightoni was subjected to micro-tomographic analysis at the Museum für Naturkunde Berlin, using a Phoenix nanotomX-ray|s tube. The cone-beam reconstruction was performed using the datos|x-reconstruction software (GE Sensing & Inspection Technologies GMBH phoenix|x-raydatos|x 2.0) and the data were visualised in VGStudio Max 2.2. Teeth (including empty sockets) were counted on all dentigerous bones.
Museum specimen number prefixes
Results
Molecular phylogenetics
The inferred phylogenies are broadly congruent with those presented by

ML phylogeny showing relationships of the newly sampled Boiga (in blue) and sequences of other available congeners. ML bootstrap support and BI posterior probability support = />75 or 0.75 is shown at each internal branch. Holotype of B. whitakeri (
The uncorrected pairwise genetic distance between the two samples of B. dightoni and the holotype of B. whitakeri, is 0.9–1.2% and 0.2–0.4% in cyt b and 16S, respectively (Table
Pairwise genetic distances (%) between the Boiga spp. from the Western Ghats, Eastern Ghats and Sri Lanka for both mitochondrial 16S and cyt b genes.
| CYT B | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | |
| 1 |
Boiga “whitakeri” |
|||||||||||||||||
| 2 |
Boiga dightoni |
1.2 | ||||||||||||||||
| 3 |
Boiga dightoni |
0.9 | 1.2 | |||||||||||||||
| 4 | Boiga ceylonensis | 5.1 | 5.2 | 5.0 | ||||||||||||||
| 5 |
Boiga nuchalis |
5.3 | 5.3 | 4.3 | 6.4 | |||||||||||||
| 6 | Boiga nuchalis CESS_192 | 5.2 | 5.3 | 4.3 | 6.3 | 0.4 | ||||||||||||
| 7 | Boiga nuchalis CESS_081 | 5.7 | 5.3 | 4.6 | 6.0 | 1.6 | 1.9 | |||||||||||
| 8 | Boiga nuchalis CESS_003 | 5.4 | 5.9 | 5.0 | 6.5 | 1.6 | 1.4 | 2.3 | ||||||||||
| 9 | Boiga barnesii | 13.3 | 13.2 | 12.8 | 14.2 | 13.6 | 13.0 | 13.5 | 13.2 | |||||||||
| 10 | Boiga beddomei CESS_444 | 8.9 | 8.6 | 7.9 | 9.4 | 8.7 | 8.1 | 8.5 | 8.5 | 14.0 | ||||||||
| 11 | Boiga beddomei CESS_418 | 9.3 | 9.1 | 8.3 | 9.4 | 8.4 | 7.7 | 8.3 | 8.1 | 13.7 | 2.4 | |||||||
| 12 | Boiga cf. ranawanei | 14.5 | 15.0 | 14.8 | 14.8 | 15.1 | 14.3 | 14.8 | 14.2 | 14.8 | 13.4 | 13.7 | ||||||
| 13 | Boiga flaviviridis | 13.2 | 13.7 | 13.3 | 13.5 | 14.0 | 12.9 | 12.6 | 13.3 | 16.1 | 13.2 | 13.3 | 9.3 | |||||
| 14 | Boiga flaviviridis CESS_529 | 13.4 | 13.0 | 12.6 | 13.1 | 13.3 | 12.9 | 12.9 | 13.2 | 15.7 | 13.5 | 13.3 | 9.1 | 2.2 | ||||
| 15 | Boiga thackerayi CESS_271 | 13.9 | 14.3 | 13.7 | 14.9 | 13.8 | 13.2 | 13.1 | 13.5 | 14.5 | 13.5 | 13.3 | 11.1 | 13.1 | 12.5 | |||
| 16 | Boiga thackerayi CESS_443 | 13.6 | 13.8 | 13.4 | 14.6 | 13.7 | 13.1 | 13.0 | 13.4 | 14.6 | 13.1 | 13.1 | 10.8 | 12.7 | 12.2 | 0.6 | ||
| 17 | Boiga thackerayi CESS_292 | 14.2 | 14.2 | 14.1 | 14.6 | 14.3 | 13.6 | 13.3 | 14.1 | 14.6 | 13.8 | 14.1 | 11.1 | 12.7 | 12.0 | 3.6 | 3.3 | |
| 18 | Boiga thackerayi BNHS_2371 | 13.6 | 14.1 | 13.8 | 14.6 | 14.0 | 13.1 | 13.0 | 13.4 | 14.7 | 13.1 | 13.1 | 10.5 | 12.5 | 12.2 | 0.6 | 0.0 | 3.3 |
| 16S | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||||
| 1 |
Boiga “whitakeri” |
|||||||||||||||||
| 2 |
Boiga dightoni |
0.2 | ||||||||||||||||
| 3 |
Boiga dightoni |
0.4 | 0.2 | |||||||||||||||
| 4 | Boiga ceylonensis | 0.8 | 0.6 | 0.4 | ||||||||||||||
| 5 | Boiga nuchalis CESS_003 | 0.7 | 0.9 | 0.7 | 1.1 | |||||||||||||
| 6 | Boiga nuchalis CESS_081 | 0.6 | 0.8 | 0.6 | 1.0 | 0.0 | ||||||||||||
| 7 | Boiga nuchalis CESS_192 | 0.6 | 0.8 | 0.6 | 1.0 | 0.0 | 0.0 | |||||||||||
| 8 | Boiga barnesii | 4.0 | 3.7 | 4.0 | 4.0 | 4.6 | 4.6 | 4.6 | ||||||||||
| 9 | Boiga beddomei CESS_418 | 1.5 | 1.7 | 1.9 | 2.3 | 1.8 | 1.7 | 1.7 | 3.3 | |||||||||
| 10 | Boiga beddomei CESS_444 | 1.7 | 1.9 | 2.1 | 2.5 | 2.4 | 2.3 | 2.3 | 3.8 | 0.8 | ||||||||
| 11 | Boiga cf. ranawanei | 3.6 | 3.8 | 3.6 | 4.0 | 3.5 | 3.4 | 3.4 | 4.6 | 3.1 | 4.0 | |||||||
| 12 | Boiga flaviviridis CESS_529 | 2.9 | 2.7 | 2.5 | 2.5 | 3.1 | 3.1 | 3.1 | 3.8 | 3.4 | 3.8 | 2.7 | ||||||
| 13 | Boiga thackerayi CESS_292 | 1.9 | 2.1 | 2.3 | 2.3 | 2.6 | 2.5 | 2.5 | 3.5 | 2.1 | 2.5 | 3.4 | 2.9 | |||||
| 14 | Boiga thackerayi CESS_443 | 2.7 | 2.5 | 2.7 | 2.7 | 3.5 | 3.4 | 3.4 | 3.1 | 2.5 | 2.9 | 3.6 | 3.4 | 0.8 | ||||
| 15 | Boiga thackerayi CESS_271 | 2.7 | 3.1 | 3.4 | 3.4 | 3.5 | 3.4 | 3.4 | 3.8 | 2.5 | 2.9 | 3.4 | 3.6 | 0.8 | 0.2 | |||
Morphological comparison of B. dightoni and B. whitakeri
Both specimens of Boiga sp. (
Meristic and morphometric data (in mm) for Boiga dightoni and Boiga nuchalis examined in this study.
| Species | B. dightoni | B. dightoni | B. dightoni | B. dightoni | B. dightoni | B. dightoni | B. dightoni | B. nuchalis | B. nuchalis | B. nuchalis | B. nuchalis | B. nuchalis | B. nuchalis |
| Voucher |
|
|
|
|
|
|
1940.10.13.19 | 74.4.29.935 | 74.4.29.934 | 74.4.29.933 | 74.4.29.936 |
|
|
| Location | Arippa, Kerala | Palagapandy, Kerala | Peermed, Kerala | Topslip, Tamil Nadu | Travancore | Ponmudi, Kerala | Kottayam, Kerala | Malabar, Western Ghats | Malabar, Western Ghats | Malabar, Western Ghats | Malabar, Western Ghats | Yercaud, Tamil Nadu | Wayanad, Kerala |
| Sex | Female | Unsexed | Male | Male | Female | Male | Female | Male | Female | Male | Juvenile Female | Female | Female |
| HL | 22.46 | 21.8 | 27.9 | 21.56 | 26.2 | NA | 24 | 20.3 | 21.2 | 25.5 | 12.3 | 16.64 | 15.5 |
| HW | 13.1 | 15.8 | 16.3 | 13.51 | 15.8 | NA | 14.2 | 10.79 | 13.9 | 15.85 | 7.3 | 12.65 | 10.6 |
| HH | 8.1 | 9.1 | 9.58 | 9.37 | 10 | NA | 9.2 | 6.7 | 9.4 | 10.5 | 4.5 | 6.31 | 5.4 |
| FL | 5.3 | 5.7 | 6.16 | 6.2 | 6 | NA | 5.83 | 5.2 | 5 | 5.94 | 3.96 | 4.22 | 4.5 |
| FW | 4.9 | 5.4 | 5.19 | 5.1 | 5.6 | NA | 5.9 | 4.4 | 4.1 | 5.4 | 2.85 | 3.45 | 3.8 |
| FrSN | 4.7 | 5 | 6.4 | 4.86 | 6.5 | NA | 6.05 | 5.2 | 5.15 | 6.73 | 2.85 | 4.2 | 3.5 |
| E-S | 5.8 | 6.3 | 7.6 | 5.7 | NA | NA | NA | NA | NA | NA | NA | 4.8 | 4.3 |
| E-N | 4.1 | 3.9 | 4.5 | 3.3 | NA | NA | NA | NA | NA | NA | NA | 3.1 | 2.7 |
| ED | 3.9 | 4.3 | 4.6 | 4.2 | NA | NA | NA | NA | NA | NA | NA | 3.2 | 3 |
| IO | 7.1 | NA | 9.3 | 7.4 | NA | NA | NA | NA | NA | NA | NA | 5.6 | 5.1 |
| SVL | 824 | 832 | 1000 | 780 | 935 | 932 | 760 | 740 | 722 | 1010 | 335 | 586 | 465 |
| TL | 206 | 92 | 268 | 203 | 234 | 245 | 191* | 200 | 181 | 255* | 80 | 157 | 120 |
| Bands on body | not visible | 80 | 76 | 80 | not visible | NA | 65 | 71 | 85 | 80 | 84 | 98 | 52 Visible |
| Bands on tail | not visible | 18+ | 28 | not visible | not visible | NA | 10 | 16 | 21 | 25 | 33 | 22-26 | not visible |
| Preoculars | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 |
| Postoculars | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 | 2,2 |
| Supralabials | 8,8 | 8,8 | 8,8 | 8,8 | 8,8 | 8,8 | 8,8 | 8,8 | 9,8 | 8,8 | 8,8 | 8,8 | 8,8 |
| Infralabials | 11,11 | 11,11 | 11,11 | 11,11 | 12,11 | 13,12 | 12,12 | 11,11 | 12,11 | 11,11 | 11,11 | 11,11 | 11,11 |
| Temporals | 2+4/2+3 | 2+4/2+3 | 3+4/2+3 | 3+4/2+4 | 2+3/2+3 | 4+4/3+4 | 3+3/3+3 | 3+3/3+3 | 3+4/3+4 | 2+2/2+3 | 2+4/2+4 | 3+4/3+4 | 3+3/3+4 |
| Preventrals | 2 | 1 | 2 | 2 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | ||
| Ventrals | 246 | 249 | 239 | 239 | 241 | 248 | 229 | 242 | 233 | 240 | 244 | 239 | 230 |
| Subcaudals | 76+ | 110 | 103/104 | 102 | 99 | 112 | 87* | 104 | 95 | 99* | 102 | 98 | 100 |
| Anal | single | single | single | single | single | single | single | single | single | single | single | single | single |
| DSR | 23:23:17 | 23:23:17 | 23:23:17 | 23:23:17 | 23:23:17 | 21/23/15 | 23:23:17 | 21:21:15 | 21:21:15 | 21:21:15 | 21:21:15 | 23:21:15 | 23:21:15 |
| * indicates an incomplete tail. | |||||||||||||
Dorsal scale row reduction formulae for some of the Boiga specimens examined in this study presented below. *between ventral 9 and 14 counts are not possible because of the damaged vertebral region. Additionally, there are several reductions and additions of the paravertebral scale row between the corresponding ventrals of 194 to 209. *** dorsal scales damaged and is not possible to find the area of scale reduction:
Boiga dightoni

———————————————————————————————————————————————
Boiga “whitakeri”
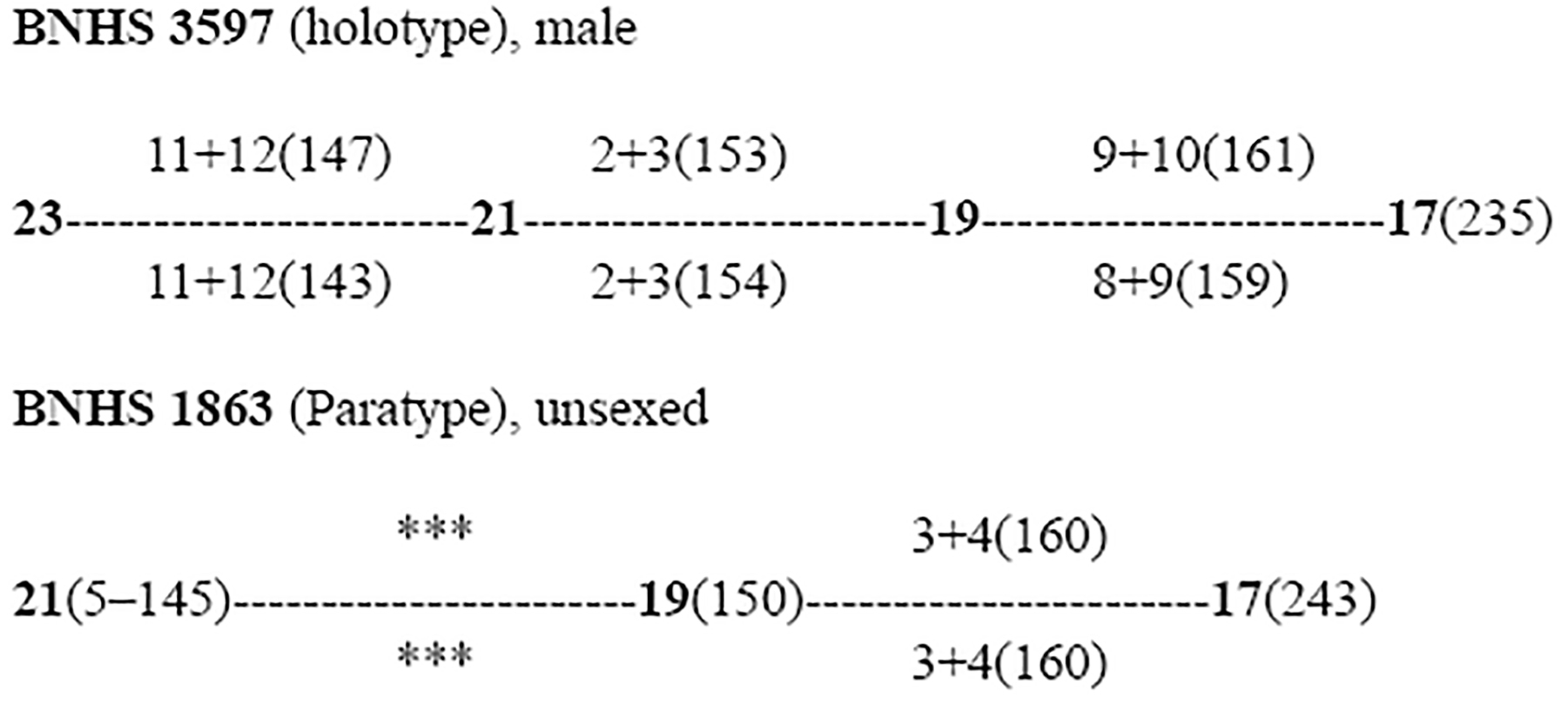
———————————————————————————————————————————————
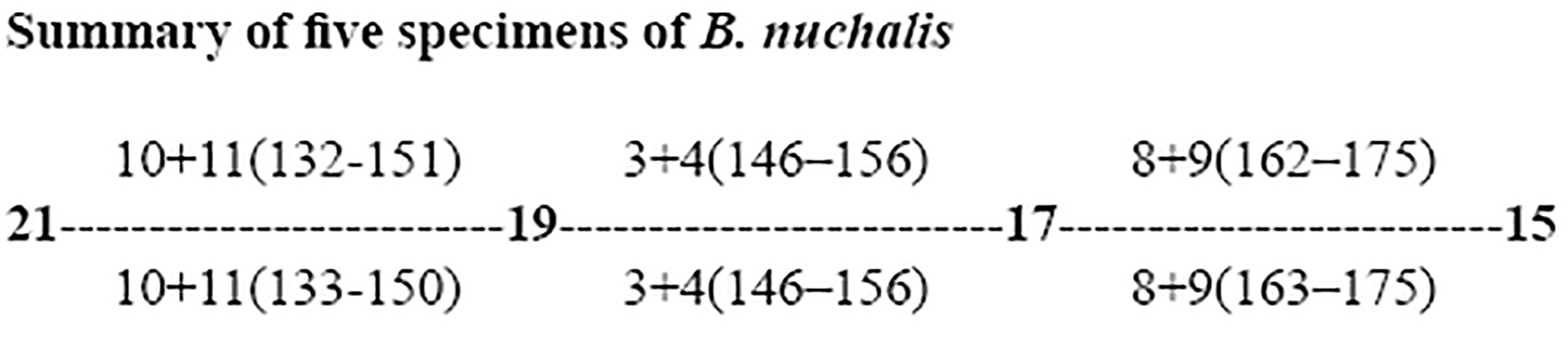
Boiga dightoni
Dipsas dightoni
Dipsadomorphus dightoni – Boulenger 1896, p. 69.
Boiga dightoni
–
Boiga whitakeri Ganesh, Mallik, Achyuthan, Shanker & Vogel, 2021 p. 453, fig. 3, syn. nov.
Taxonomic comments
A detailed description of the external morphology of the holotype of Boiga dightoni (
Based on the morphological data from the two specimens collected during this study, including the specimen from the type locality (Peermed, Kerala) of Boiga dightoni, we confidently identify these two specimens as B. dightoni. Our morphological examination of the types and non-type materials of Boiga whitakeri, B. dightoni and B. nuchalis provide evidence that led us to conclude that the holotype of B. whitakeri is conspecific with B. dightoni. This is consistent with our molecular analyses, in which the holotype of B. whitakeri is nested within the samples (including the topotype) that we identify as B. dightoni. On the other hand, the type series (
Morphology
A medium-sized Boiga (greatest TL 1000 mm (male), 935 mm (female)); 229–249 ventrals, 99–112 divided subcaudals; 13/14 teeth on maxilla and 7 on palatine; dorsal scales smooth, 23:23:19 in rows; dorsal scale reduction from 23 to 21 rows occurs between ventrals 123–144 and the reduction from 21 to 19 occurs between ventrals 148–155. Dorsum reddish dun to olive greenish with dorsal light brown to dark bands. Head with dark marking dorsally (rarely absent) and a dark laterocular stripe present.
Colouration in life and preservative
Based on the (live and museum) specimens examined and information available from the literature, we report two different colour morphs in B. dightoni.
Morph 1 (n = 5)
Reddish dun-coloured dorsum with faint reddish bands on the body (rarely absent) with or without distinct dark marking on the head, and ventral scales uniformly creamish white (Figs


Morph 2 (n = 5)
Olive greenish dorsum with black bands (76–80) on the body, with distinct marks on the head and a postocular stripe that ends shortly behind the fissure of the mouth, and irregular small dark blotches along the paraventral scales (Figs


Based on the specimens examined here, it is also clear that these two colour morphs are not explained by sexual dichromatism because both male and female specimens are known for both morphs. For example, the male specimens
Description of hemipenis of ZSI-CZRC -V-7541 (Fig. 7 )
The right hemipenis is fully everted and removed in situ for further analysis. The hemipenis is sub-cylindrical and moderately elongate (length: 17.0 mm, maximum width: 5.7 mm), extending to the 7th subcaudal. The sulcus is undivided, bounded by thick walls on both sides, and terminates at the centre of the lobe. It can be differentiated into three zones; the proximal zone is covered with 4–6 rows of spines (~40% of the total length), the middle zone with 5 or 6 rows of spinulate flounces arranged transversely (~35% of the total length), and the distal calyculate area (~25%) with 4 or 5 rows of irregular calyces with papilate edges. The sulcus spermaticus is exposed before entering the calyculate area. There is not much variation in the arrangements of spines and body calyces on sulcate and asulcate sides. The overall structure of the hemipenis of

Dentition based on the holotype of B. dightoni (BMNH 1946.1.1.32) (left/right order)
Maxillary bone with 13/14 prediastemal teeth, followed by a distinct diastema that is as long as the socket of the last prediastemal tooth and followed by two distinctly enlarged, grooved and posteriorly bent postdiastemal teeth. Prediastemal teeth increase in size posteriorly, the anterior three distinctly posteriorly hooked, the following with less pronounced curvature. On the left side, prediastemal teeth 1, 4, 5, 7, 9, 11, and 13 missing, maxilla broken behind the diastema. On the right side, prediastemal teeth 2–4, 6, 8, 10, 12, 13 and anterior postdiastemal tooth are missing.
Palatine bone with 7/7 posteriorly curved teeth, anterior ones as long as the middle prediastemal teeth, slightly decreasing in size posteriorly. Teeth 1, 5 and 7 are loose, and tooth 3 missing on left side. Teeth 1, 3 and 5 are loose on the right side. Lateral to each palatine tooth is a single replacement tooth at different growth stages. Pterygoid bone with 18/16 posteriorly curved teeth, first one half as long as last palatine tooth, gradually decreasing in size posteriorly, last one minute. Teeth 2, 4, 6, 8, 10, 12, and 14–16 missing on left side, teeth 2, 4, 6, 8, and 10 loose, and 11, 13, and 15 missing on right side. The posterior 45% of the pterygoid bone is without teeth.
Mandibular bone with 20/20 posteriorly curved teeth, shorter than maxillary and palatine teeth, gradually decreasing in size posteriorly. Medial to each mandibular tooth is a single replacement tooth in different growth stages. Teeth 1, 3–7, 9, 11, 13, 15–17, and 19 missing, tooth 2 loose on left side, teeth 1, 3, 5, 7, 9, 11, 13, 15, 17, and 19 missing, and tooth 2 loose on right side. Mandibular bone broken behind tooth 13 on left side.
Distribution
Based on currently available data, Boiga dightoni is widely distributed in the southern Western Ghats (south of the Palghat Gap), at elevations of 9–1258 m (Appendix
Discussion
Intraspecific colour polymorphism has been reported in several colubrid genera (
The holotype of Boiga dightoni (in preservative) is uniform in colour without dorsal markings and this probably led to several misidentifications in the past. Beyond the specimens examined here, it is probable that several individuals of B. dightoni are misidentified as B. nuchalis based on colour pattern. For example, at least two records (https://www.inaturalist.org/observations/37460080, 86841689) identified as B. nuchalis may actually represent B. dightoni. As mentioned above, only a few records of B. dightoni are available in the literature and this might be mainly because of its overall similarity with B. nuchalis, a much more commonly encountered species that partly overlaps in geographic range with the former. It is likely that, at least in some places, these two species are sympatric. Hence, we hereby caution against identifying these species solely based on the colour pattern, especially from the southern Western Ghats where both B. dightoni and B. nuchalis are present. Our results highlight the importance of careful examination of type specimens when describing new, similar and closely related species, especially in the absence of molecular data. Wherever possible, it is also preferable to select well-preserved and undamaged specimens when designating name-bearing types.
During this study, we also examined the type series of Boiga thackerayi. In the original description (
Revised key to the species in the Boiga ceylonensis complex of Western Ghats, India and Sri Lanka, modified from Ganesh et al. (2021)
| 1a | Midbody scale rows 19 | 2 |
| 1b | Midbody scale rows 21, temporal scales larger than body scales | B. nuchalis |
| 1c | Midbody scale rows 23, temporal scales subequal to body scales | B. dightoni |
| 2a | Dorsum greenish | B. flaviviridis |
| 2b | Dorsum brownish | 3 |
| 3a | Subcaudals > 110 pairs, preocular 1 | B. beddomei |
| 3b | Subcaudals > 110 pairs, preocular 2 | B. ranawanei |
| 3c | Subcaudals < 110 pairs | 4 |
| 4a | Ventrolateral white blotches absent | 5 |
| 4b | Ventrolateral white blotches present | 6 |
| 5 | Crown markings on parietals conspicuous and dark; bands dark, prominent | B. ceylonensis |
| 6a | Preocular 1; dorsum barred | B. thackerayi |
| 6b | Preoculars 3; dorsum blotched | B. barnesii |
Acknowledgements
We thank the Kerala Forest Department for permits (WL10-636/2021 dated 16/10/2021) and support.
We thank Ashok Captain for his support and advice on Indian snake taxonomy. We thank Dhanu Paran, Vinu J George, Amal Varghese and Akhil KS for their hospitality, Jishnu N, Arun Vijayakumar, Siddharth S, Joju CT, Lal V, Nihal J, Sanjay C, Vignesh B, Nithin D, Ameer K, Santhosh KT, Aravind, Amirtha Balan and Nobin Raja for their support in the field. Patrick Campbell, NHM, London for his support to DV and loans to Frank Tillack. K. A. Subramanian, Office in charge, ZSI Chennai and S. R. Ganesh, Chennai Snake Park Trust for sharing images of the specimen at ZSI, Chennai. SN thanks Kartik Shanker for access to the specimen at CES, Bangalore. SN thanks Aravind NA (Senior Fellow, ATREE) for his support at ATREE. We thank the National Geographic grant (NGS-63816R-19) for the support for fieldwork and museum visits. VD’s contribution was supported in part by the Humboldt fellowship hosted by Uwe Fritz at the Senckenberg Dresden. We thank Saunak Pal and an anonymous reviewer for their comments on the initially submitted version of this manuscript.
References
- Adalsteinsson SA, Branch WR, Trape S, Vitt LJ, Hedges SB (2009) Molecular phylogeny, classification, and biogeography of snakes of the family Leptotyphlopidae (Reptilia, Squamata). Zootaxa 2244: 1–50. https://doi.org/10.11646/zootaxa.2244.1.1
- Boulenger GA (1894) Description of a new snake found in Travancore, by Mr. S. Dighton. Pirmaad. Journal of the Bombay Natural History Society 8: 528.
- Cox CL, Davis Rabosky AR (2013) Spatial and temporal drivers of phenotypic diversity in polymorphic snakes. The American Naturalist 182: E40–E57. https://doi.org/10.1086/670988
- Dowling HG (1951a) A proposed method of expressing scale reductions in snakes. Copeia 1951: 131–134.
- Dowling HG (1951b) A proposed standard system of counting ventrals in snakes. British Journal of Herpetology 1: 97–99.
- Dowling HG, Savage JM (1960) A guide to the snake hemipenis: a survey of basic structure and systematic characteristics. Zoologica 45: 17–28.
- Ganesh SR, Achyuthan NS, Chandramouli SR, Vogel G (2020) Taxonomic revision of the Boiga ceylonensis group (Serpentes: Colubridae): re-examination of type specimens, redefinition of nominate taxa and an updated key. Zootaxa 4779: 301–332. https://doi.org/10.11646/zootaxa.4779.3.1
- Ganesh SR, Mallik AK, Achyuthan NS, Shanker K, Vogel G (2021) A new species of Boiga (Serpentes: Colubridae) from the Southern Western Ghats of India with a molecular phylogeny and expanded characterisation of related species. Zootaxa 4981: 449–468. https://doi.org/10.11646/zootaxa.4981.3.2
- Giri VB, Deepak V, Captain A, Pawar S, Tillack F (2019) A new species of Boiga Fitzinger, 1826 (Serpentes: Colubridae) from the northern Western Ghats of India. Journal of the Bombay Natural History Society 116: 1–11.
- Higgins D, Thompson J, Gibson T, Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research 22: 4673–4680. https://doi.org/10.1093/nar/22.22.4673
- Inger RF, Shaffer HB, Koshy M, Bakde R (1984) A report on a collection of amphibians and reptiles from the Ponmudi, Kerala, South India. Journal of the Bombay Natural History Society 81: 406–427.
- Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS (2017) ModelFinder: Fast model selection for accurate phylogenetic estimates. Nature Methods 14: 587–589. https://doi.org/10.1038/nmeth.4285
- Kanagavel A, Ganesh SR (2021) Recent record of the rare Travancore Catsnake, Boiga dightoni (Boulenger 1894) (Reptilia: Colubridae), from the Ponmudi Hills in the southern Western Ghats, India. Reptiles and Amphibians 28: 67–70.
- Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870–1874. https://doi.org/10.1093/molbev/msw054
- Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B (2017) PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Molecular Biology and Evolution 34: 772–773. https://doi.org/10.1093/molbev/msw260
- Mohapatra PP, Das A, Tillack F, Dutta SK (2009) Taxonomy, natural history, and distribution of Boiga forsteni (Duméril, Bibron et Duméril, 1854) (serpentes: Colubridae) from Orissa, India. Russian Journal Herpetology 16: 243–252.
- Murthy TSN (1984) A record of the rare cat snake, Boiga dightoni (Boulenger) (Serpentes: Colubridae). The Snake 17: 84.
- Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ (2015) IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Molecular Biology and Evolution 32: 268–274. https://doi.org/10.1093/molbev/msu300
- Palumbi SR, Martin AP, Romano SL, McMillan WO, Stice L, Grabowski G (1991) The Simple Fool’s Guide to PCR. Version 2. University of Hawaii, Honolulu, 43 pp.
- Palacios-Aguilar R, Colín-Martínez VH, Hernández-Rubio S, Canseco-Márquez L, Nieto-Montes de Oca A, Ochoa-Ochoa LM (2021) Another case of colour pattern polymorphism in earth snakes of the genus Geophis (Dipsadidae) from southern Mexico. Journal of Natural History 55: 2985–2997. https://doi.org/10.1080/00222933.2021.2024906
- Pavón-Vázquez CJ, García-Vázquez UO, Blancas-Hernández JC, de Oca ANM (2011) A new species of the Geophis sieboldi group (Squamata: Colubridae) exhibiting color pattern polymorphism from Guerrero, Mexico. Herpetologica 67: 332–343.
- Ronquist F, Teslenko M, Mark VDP, Ayres D, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. https://doi.org/10.1093/sysbio/sys029
- Russell P (1796) An account of Indian serpents, collected on the Coast of Coromandel; containing descriptions and drawings of each species; together with experiments and remarks on their several poisons. George Nicol, London, vii+91 pp., 46 pls.
- Sabaj MH (2020) Codes for Natural History Collections in Ichthyology and Herpetology. Copeia 108: 593–669. https://doi.org/10.1643/ASIHCODONS2020
- Samarawickrama VAMPK, Samarawickrama VAP, Wijesena NM, Orlov NL (2005) A new species of genus Boiga (Serpentes: Colubridae: Colubrinae) from Sri Lanka. Russian Journal Herpetology 12: 213–222.
- Smith MA (1943) The Fauna of British India, Ceylon and Burma, Including the Whole of the Indo-Chinese Sub-Region. Reptilia and Amphibia. 3 (Serpentes). Taylor and Francis, London, 583 pp.
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA 5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Molecular Biology and Evolution 28: 2731–2739. https://doi.org/10.1093/molbev/msr121
- Tillack F, Narayanan S, Deepak V (2021) On the identity, nomenclatural status and authorship of Coluber monticolus Cantor, 1839 (Reptilia: Serpentes). Zootaxa 4990: 134–146. https://doi.org/10.11646/zootaxa.4990.1.8
- Trifinopoulos J, Nguyen LT, von Haeseler A, Minh BQ (2016) W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Research 44: 232–235. https://doi.org/10.1093/nar/gkw256
- Uetz P, Freed P, Hošek J (2022) The Reptile Database. Available from: http://reptile-database.reptarium.cz (accessed 31 August 2021)
- van Rooijen J, Wood PL, Grismer JL, Grismer LL, Grossmann W (2011) Color pattern dimorphism in the colubrid snake Oligodon purpurascens (Schlegel, 1837) (Reptilia: Squamata). Russian Journal of Herpetology 18: 215–220.
- Wallach V, Williams KL, Boundy J (2014) Snakes of the World: A catalogue of living and extinct Species. CRC Press,Taylor & Francis Group, London, xxvii + 1209 pp.
- Weinell JL, Barley AJ, Siler CD, Orlov NL, Ananjeva NB, Oaks JR, Burbrink FT, Brown RM (2021) Phylogenetic relationships and biogeographic range evolution in cat-eyed snakes, Boiga (Serpentes: Colubridae). Zoological Journal of the Linnean Society 192: 169–184. https://doi.org/10.1093/zoolinnean/zlaa090
Appendix 1
Genbank voucher numbers for the samples used in this study. New sequences generated for this study are marked in bold.
| Species | Voucher no. | Location | CYT B | 16S |
| Boiga barnesii | RAP0452 | Sri Lanka | KC347469 | KC347345 |
| Boiga beddomei | CESS 418 | Mhadei WLS, Goa | MT733292 | MT734906 |
| Boiga beddomei | CESS 444 | Mahabaleswar, Maharashtra, India | MT733294 | MT734908 |
| Boiga bourreti | ZISP 32786 | Mang Canh, Kon Plong, Kon Tum, Vietnam | MN962356 | – |
| Boiga ceylonensis | RS-Y | Sri Lanka | KC347467 | KC347347 |
| Boiga cf. ranawanei | RAP0450 | Sri Lanka | KC347466 | KC347346 |
| Boiga cyanea | CHS553 | — | MK201410 | MK194064 |
| Boiga cynodon | — | Palawan Islands, Philippines | KC010340 | AF139566 |
| Boiga dendrophila | — | — | AF471089 | – |
| Boiga dightoni | BNHS 3597 | DevarMalai, Tamil Nadu, India | MT733284 | MT734897 |
| Boiga dightoni | ZSI-CZRC-V-7541 | Peermed, Kerala, India | OP948298 | OP955936 |
| Boiga dightoni | BNHS 3617 | Arippa, Kerala, India | OP948299 | OP955937 |
| Boiga drapiezii | LSUHC7295 | — | KX660482 | KX660210 |
| Boiga flaviviridis | — | Meghamalai, Tamil Nadu, India | MN508360 | – |
| Boiga flaviviridis | CESS 529 | Horsley hills, Andhra Pradesh, India | MT733297 | MT734911 |
| Boiga forsteni | RAP0540 | Sri Lanka | KC347468 | KC347348 |
| Boiga irregularis | — | — | FJ710794 | AF139551 |
| Boiga jaspidea | LSUHC7656 | Endau-Rompin, Johor, West Malaysia | KX660484 | KX660212 |
| Boiga kraepelini | CHS115 | — | MK201272 | MK193920 |
| Boiga multomaculata | CHS760 | — | MK201511 | MK194200 |
| Boiga nigriceps | LSUHC7020 | — | KX660485 | KX660213 |
| Boiga nuchalis | BNHS 3618 | Yercaud, Tamil Nadu, India | OP948300 | – |
| Boiga nuchalis | CESS 003 | Coorg, Karnataka, India | MT733270 | MT734883 |
| Boiga nuchalis | CESS 081 | Meppadi,Wynad, Kerala, India | MT733274 | MT734887 |
| Boiga nuchalis | CESS 192 | Kolli Hills, Tamil Nadu, India | MT733282 | MT734895 |
| Boiga ochracea | CAS215390 | Yinpaungtaing Village, Yin Ma Bin Township, Sagaing, Myanmar | MN962367 | – |
| Boiga quincunciata | CAS221434 | Putao Dist. Myanmar | KX660451 | KX660177 |
| Boiga schultzei | KU 327776 | Estrella Falls Park, Estrella, Narra, Palawan,Philippines | MN962368 | – |
| Boiga siamensis | LSUHC8502 | O’lakmeas, Pursat Province, Cambodia | KX660487 | KX660215 |
| Boiga thackerayi | CESS_ 271 | Thadiyandamol, Karnataka, India | MT733286 | MT734899 |
| Boiga thackerayi | BNHS 2371 | Koyna, Maharashtra, India | MN508359 | – |
| Boiga thackerayi | CESS 292 | KalakadMundanthurai Tiger Reserve, Tamil Nadu, India | MT733287 | MT734900 |
| Boiga thackerayi | CESS 443 | Mahabaleswar, Maharashtra, India | MT733293 | MT734907 |
| Boiga trigonata | RS-143 | Sri Lanka | KC347475 | KC347349 |
| Boiga westermanni | — | India | MG428713 | MG428711 |
| Telescopus tripolitanus | BEV9377 | Mauritania | JX315531 | MK372141 |
| Telescopus variegatus | — | — | MK373093 | MK372142 |
| Toxicodryas pulverulenta | CAS220642 | — | KX660460 | KX660187 |
Appendix 2
List of Boiga specimens examined in this study. Specimens examined for scale reductions are marked in bold.
Boiga dightoni (n = 9)
Morph 1.
Morph 2.
Boiga beddomei (n = 1).
Boiga ceylonensis (n = 7).
Boiga flaviviridis (n = 1).
Boiga nuchalis (n = 10).
Boiga thackerayi (n = 4).
Appendix 3

Appendix 4
Gazetteer of confirmed locality records for Boiga dightoni and B. nuchalis in India. Localities where we verified only images for confirmation are marked with an asterisk.
| Species | Current locality | Latitude | Longitude |
| Boiga nuchalis* | Bekkinjaddi, Audala, Karnataka, India | 14.73401 | 74.75821 |
| Boiga nuchalis* | Guddekeri, Karnataka, India | 13.56631 | 75.1342 |
| Boiga nuchalis* | Honnavar, Karnataka, India | 14.27975 | 74.44393 |
| Boiga nuchalis* | Magod Falls, Karnataka, India | 14.86487 | 74.75922 |
| Boiga nuchalis | Mavinagudi, Karnataka, India | 14.92432 | 74.82812 |
| Boiga nuchalis* | Guddekeri, Karnataka, India | 13.56631 | 75.1342 |
| Boiga nuchalis* | Mayfield, Tamil Nadu, India | 11.55756 | 76.43534 |
| Boiga nuchalis* | Rockwood Estate, Tamil Nadu, India | 11.53503 | 76.40159 |
| Boiga nuchalis* | Hope Estate, Tamil Nadu, India | 11.58956 | 76.06355 |
| Boiga nuchalis* | Pilloor, Tamil Nadu, India | 11.30443 | 76.80602 |
| Boiga nuchalis* | Adderly Estate, Nilgiris, Tamil Nadu, India | 11.35901 | 76.85699 |
| Boiga nuchalis | Kolli hills, Tamil Nadu, India | 11.295 | 78.377 |
| Boiga nuchalis | Yercaud, Shervaroys, Tamil Nadu, India | 11.83435 | 78.24079 |
| Boiga nuchalis | Sirumalai hills, Tamil Nadu, India | 10.20065 | 77.99901 |
| Boiga nuchalis* | Shimoga, Karnataka, India | 13.51287 | 75.14139 |
| Boiga nuchalis* | Yevakapadi, Karnataka, India | 12.20958 | 75.64052 |
| Boiga nuchalis | Kasargod, Kerala, India | 12.49293 | 75.27597 |
| Boiga nuchalis | Coorg, Karnataka, India | 12.20958 | 75.64052 |
| Boiga nuchalis | Attakatti, Anamalai Tiger Reserve, Tamil Nadu, India | 10.44754 | 76.9861 |
| Boiga nuchalis | Mannarkad, Kerala, India | 11.05046 | 76.47072 |
| Boiga nuchalis | Vythiri, Wayanad, Kerala, India | 11.51478 | 76.03951 |
| Boiga nuchalis | Siruvani, Tamil Nadu, India | 10.987 | 76.622 |
| Boiga nuchalis | Vazhachal, Kerala, India | 10.303 | 76.593 |
| Boiga nuchalis* | Chimmini dam road, Kerala, India | 10.43104 | 76.49101 |
| Boiga nuchalis | Taliparamba, Kerala, India | 12.022472 | 75.363804 |
| Boiga nuchalis* | Kervashe Village, Karnataka, India | 13.258421 | 75.081153 |
| Boiga nuchalis | Kalpetta, Wyanad, Kerala, India | 11.588 | 76.1 |
| Boiga nuchalis | Pullompara, Kerala, India | 10.086 | 76.511 |
| Boiga nuchalis | Iruppu falls, Kerala, India | 11.969 | 75.985 |
| Boiga nuchalis | Thadiyendamol, Karnataka, India | 12.229 | 75.623 |
| Boiga nuchalis* | Potachipara, Bramagiri, Karnataka, India | 12.077 | 75.805 |
| Boiga nuchalis | Forests of west coast of Malabar, Kerala, India | 11.545426 | 75.757901 |
| Boiga dightoni | Peermade, Kerala | 9.576675 | 77.03061 |
| Boiga dightoni | Aanapara, Ponmudi hills | 8.69 | 77.1 |
| Boiga dightoni | Ponmudi, Kerala, India | 8.752984 | 77.12104 |
| Boiga dightoni | Devermala, Kerala, India | 9.173 | 77.261 |
| Boiga dightoni | Arippa, Kerala, India, Kerala, India | 8.832483 | 77.03245 |
| Boiga dightoni* | Coutrallam, Tamil Nadu, India | 8.923837 | 77.25514 |
| Boiga dightoni | Kottayam, Kerala, India | 9.579248 | 76.54887 |
| Boiga dightoni | Topslip, Tamil Nadu, India | 10.46901 | 76.84185 |
| Boiga dightoni | Manampalli, Anamalai Tiger Reserve, Tamil Nadu, India | 10.35406 | 76.87829 |
| Boiga dightoni* | Kalakad Mundanthurai Tiger Reserve, Tamil Nadu, India | 8.880428 | 77.28482 |
| Boiga dightoni | Palagapandy, Nelliampathy, Kerala, India | 10.56112 | 76.7304 |
Appendix 5
Scale reduction formula for the two Boiga thackerayi type specimens at