Research Article |
|
Corresponding author: V. Deepak ( veerappandeepak@gmail.com ) Academic editor: Uwe Fritz
© 2021 V. Deepak, Surya Narayanan, Pratyush P. Mohapatra, Sushil K. Dutta, Gnanaselvan Melvinselvan, Ashaharraza Khan, Kristin Mahlow, Frank Tillack.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Deepak V, Narayanan S, Mohapatra PP, Dutta SK, Melvinselvan G, Khan A, Mahlow K, Tillack F (2021) Revealing two centuries of confusion: new insights on nomenclature and systematic position of Argyrogena fasciolata (Shaw, 1802) (auctt.), with description of a new species from India (Reptilia: Squamata: Colubridae). Vertebrate Zoology 71: 253-316. https://doi.org/10.3897/vz.71.e64345
|
Abstract
Coluber fasciolatus Shaw, 1802 (today Argyrogena fasciolata auctt.) is the name used for a widespread terrestrial colubrid snake species inhabiting subtropical and tropical dry deciduous/thorn forests of South Asia from Pakistan to India, with unconfirmed records of distribution in Nepal, Bangladesh and Myanmar and a single doubtful record from the northern tip of Sri Lanka.
During the past 200 years, A. fasciolata (common name Banded racer) has been placed in different genera, i.e. Tyria Fitzinger, 1826, Zamenis Wagler, 1830, Coryphodon Duméril, Bibron and Duméril, 1854, and Coluber Linnaeus, 1758 where it primarly remained until the mid 1960s and exceptionally until the year 2011. Three subsequently introduced names, viz. Coluber hebe Daudin, 1803, Coluber curvirostris Cantor, 1839, and Argyrogena rostrata Werner, 1924 were synonymized with C. fasciolatus shortly after its description.
Based on a combination of characters including body pattern, external morphology and osteological differences
Specimens of Banded racer found in the southeastern part of peninsular India are morphologically and genetically distinct from populations of the rest of the distribution area. These populations from central and southern Tamil Nadu state represent a different species, consequently described as a new species herein. Furthermore, examination of specimens of the Banded racer from different populations across its entire range, including the type specimen of the genus Argyrogena (A. rostrata Werner), reveals a similarity in morphology with the genus Platyceps Blyth, 1860. This was further supported by molecular data which demonstrates that the genus Argyrogena is nested within Platyceps.
Keywords
Distribution, Lycodon fasciolatus comb. nov., morphology, Platyceps josephi sp. nov., Platyceps plinii comb. nov., Serpentes, South Asia, systematics
Introduction
The Banded racer, presently named Argyrogena fasciolata (auctt.), is a colubrid snake species with an assumed wide but scattered distribution in the Indian subcontinent and has an unstable taxonomic history. This species was first mentioned by the Scottish surgeon and naturalist Patrick
Our critical analysis of the original source (
Materials and methods
Sampling
We generated DNA sequence data for eight specimens of Argyrogena (auctt.) collected from different parts of peninsular India (Appendix
Molecular analysis
Genomic DNA was extracted from liver or tail tissue samples stored in absolute ethanol at –20°C. DNeasy (Qiagen) blood and tissue kits were used to extract DNA. We amplified partial sequences of two nuclear (nu) and three mitochondrial (mt) genes. Cytochrome b (cytb) and NADH dehydrogenase subunit 4 (ND4), 16S rRNA (16S) and the nu markers are oocyte maturation factor (cmos) and recombination activating gene 1 (rag1). PCR conditions followed previously reported protocols for the respective primer pairs (cytb, primers GluDG L:
Apart from the five genes for which we generated data, we also included an additional gene 12S because sequences of this marker were available for 55% of the samples. Taxon sampling was based on the recent publications on global snake and old world racer phylogenies (
All phylogenetic analyses were carried out using the CIPRES Science Gateway portal v3.3 (
Morphology
In addition to the type series and referred material (n=11) of the new species described below, comparative material examined directly includes the holotype of Argyrogena rostrata (type species of the genus Argyrogena Werner, 1924) and additional preserved museum specimens so far determined as A. fasciolata (n=50) and representative specimens from the genera Lycodon, Hemorrhois, Platyceps and Spalerosophis (n=33). Examined material is listed in Appendix
Total length, snout-vent length and tail length were measured with thread and a ruler to the nearest 1 mm. Other dimensions were recorded with dial callipers, to the nearest 0.1 mm. Identification of sex was performed by dissection (presence or absence of hemipenes, inspected through a subcaudal incision at the tail base).
The hemipenis of freshly euthanized specimens were everted by pressing the organ until it was everted to the maximum extent, after which the retractor muscle was cut at the level of the 10th subcaudal scale. From preserved specimens in which the hemipenis was not everted, it was removed through a longitudinal incision at 15th subcaudal and detached from the body by cutting the retractor muscle. The organ was separated by making an incision around its circumference at the cloacal region and was immersed in warm water (60°C) for about 5 minutes to soften the tissue. It was then slowly everted with a blunt-end forceps by gently pushing the organ from the distal to proximal end. Afterwards, the hemipenis was softened by again immersing it in warm water for intervals of 5–10 minutes. While everting, petroleum jelly was applied to the inner edge for smooth sliding of the inner ornamented layer. Care was also taken to avoid puncturing the hemipenial wall due to the rubbing of the spines. After eversion, the organ was inflated with petroleum jelly and tied at the base with a thread. Later it was immersed in 1% alizarin red solution in 70% ethanol and removed from the solution after one hour of staining. The calcified spicules were stained using this process and excess stain was removed by immersion of the hemipenis in 70% alcohol for approximately 30 minutes. Detailed photographs of the stained hemipenes were taken using Nikon D 5100 with a 100 mm macro lens (Nikon). Descriptions of hemipenial characters and terminology follow
Length and width of head scales were measured at the longest and widest points of the respective scale(s). Interocular width was measured on top of the head at the shortest distance between the eyes. The number of ventral scales was counted according to
To evaluate character states of midbody vertebrae we created µCT-scans of postcranial skeletons for six “Banded racers”, four Platyceps rhodorachis and one each for P. florulentus, P. ventromaculatus and the new species, respectively and also included literature data for these taxa provided by
To obtain counts of teeth by a non-invasive procedure, heads of Platyceps species and comparative material were subjected to micro-tomographic analyses with GE Inspections Technologies computer tomographs, e.g. a Phoenix nanotom X-ray|s, at the Museum für Naturkunde Berlin, Germany, and a Phoenix v|tome|x M at Chakan, Pune, India. For body scans we used a FF35 (YXLON International, Hamburg, Germany) at the Museum für Naturkunde Berlin, Germany. The scans were performed with 60–95 kV and 135–1000 μA, generating 1000 or 1440 projections with 750 or 1000 ms per scan. The different kV and µA-settings depended on the respective specimen size. Effective voxel size, i.e. resolution in three-dimensional space, ranged from 10–71.16 μm. The cone beam reconstruction was performed using the datos|x-reconstruction software (GE Sensing and Inspection Technologies GMBH phoenix|x-ray datos|x 2.1) and the data were visualized in VGStudio Max 3.0.
Abbreviations of morphological characters used in text and tables are as follows: Measures and ratios: SVL – snout-vent length; TaL – tail length; TL – total length (SVL+TaL); TaL/TL – ratio tail length/total length; HW – head width, measured at its widest lateral extension; HL – head length, measured from tip of snout to posterior edge of mandible; ED – eye diameter, measured horizontally; RW – rostral width; FL – frontal length; FW – frontal width; PrefL – prefrontal length; InL – internasal length; E-S – distance from anterior border of the eye to tip of snout; ParL – parietal length; ParW – parietal width; DSR – dorsal scale rows: numbers are given at one head length behind head, at midbody (taken at half of the total number of ventral scales), and at one head length before vent, respectively; PreVen – number of preventrals; Ven – number of ventrals; TS – total number of ventral and subcaudal scales including preventrals and terminal scale; PrO – number of preoculars; PoO – number of postoculars; T – number of temporal scales; SupL – number of supralabials; SubL – number of sublabials.
The opensource software QGIS (http://www.qgis.org) was used to prepare the map. Some of the metrics used in the IUCN Red-Listing criteria (IUCN, 2012) were used to calculate the area of occupancy (AOO) and the Extent of occurrence (EOO) of the new species using the GeoCat online tool (
Results
Molecular phylogeny
Both BI and ML analysis recover a well supported monophyletic Platyceps (including Argyrogena) as sister to Hemorrhois + Spalerosophis, also with high support (ML bootstrap 100 & BI posterior probability 1; Fig.

Map showing geomorphological features in parts of the Indian subcontinent and current locality records (see Appendix

ML phylogeny showing inferred phylogenetic relationships of Old World racers. Labelled in pink colour are the sequences generated in this study, coloured blue is a sample from this study+GenBank sequence. Numbers at internal branches are ML bootstrap values. Bootstrap values below 50 are not shown.
Systematics
Lycodon fasciolatus
Coluber. –
Coluber Fasciolatus
Shaw, 1802, p. 528, 413, “Fasciolated Snake”, type locality “India”, based on
Coluber fasciolatus. –
Coluber hebe
Daudin, 1803 p. 385 ff., type locality “Coromandel”, like Coluber fasciolatus Shaw based on
[Natrix] hebe. –
Lycodon Hebe. – Boie, H. 1826, p. 238 [partim]; Boie, H. in Boie, F. 1827, col. 551 [partim];
Lycodon aulicum. – Duméril, Bibron and Duméril 1854, p. 370 [Varieté B, partim, non Lycodon aulicus (Linneus, 1758)].
Coryphodon fasciolatus. –
Lycodon anamallensis
Günther, 1864, p. 318 f., type locality “Anamallai Mountains”, holotype
Lycodon osmanhilli Taylor, 1950, p. 562 ff, pl. 20, type locality “Colombo, Ceylon”, holotype KUMNH 24141, coll. W.C. Osman-Hill.
Lycodon fasciolatus.– this work.
Coluber fasciolatus formally named by George Kearsley

Lycodon fasciolatus comb. nov. A. Holotype of Coluber fasciolatus Shaw, 1802, as well as holotype of Coluber hebe Daudin, 1803, “Nooni Paragoodoo” reproduced from
On the same basis and almost simultaneously as
In his “Catalogue of Colubrine snakes […] of the British Museum”
It is also to be noted that from the initial confusion caused by

Lycodon fasciolatus comb. nov. Previously unpublished extract from a paper sheet with glued specimens from the 1904 collection of dried snake skins preliminarily attributed to Patrick Russell showing a specimen of the “Nooni Paragoodoo” kept in the herpetological collection of the Natural History Museum, London, U.K. (
In this work, we clarify the identity of C. fasciolatus, by analysing
To determine the species affiliation we carefully compared Russell’s description and the sketch on plate 21 with published data of Lycodon species from southern and eastern India (Table
Comparison of available characters for the type specimen of C. faciolatus Shaw with other Lycodon species from South and East India. Species from India formerly allocated to the genera Dryocalamus and Dinodon are ignored. Data sourced from
| Taxon | Ventrals | Subcaudals | Preocular-frontal-contact | Collar / in contact with parietals | Colouration of upper labials | Dorsal body pattern |
| fasciolatus (holotype) | 192 | 62 | no | absent | yellow with dark brown center | short whitish bands stippled with black not extending onto ventral scale edges |
| anamallensis (Indian populations) | ♂♂ 174–186 ♀♀ 186–197 | ♂♂ 63–64 ♀♀ 59–74 | no | absent | creamy white or yellow with dark brown center | short yellowish or whitish bands stippled with black not extending onto ventral scale edges, or patternless |
| aulicus (s. str.) | ♂♂ 180–206 ♀♀ 191–215 | ♂♂ 65–72 ♀♀ 57–73 | yes | present / yes | whitish | creamy-white bands extending onto ventral scale edges |
| deccanensis | 198–214 | 66–69 | no | present / yes | light brown with irregular dark brown speckles | irregular dirty whitish bands with dark brown speckles extending onto ventral scale edges |
| fasciatus | 197–220 | 69–94 | scarcely | present / yes | dark greyish-white | dirty whitish-brown bands extending onto ventral scale edges |
| flavicollis | 210–224 | 65–72 | yes | present / usually | yellow | without or with very faint yellow bands not extending onto ventral scale edges |
| flavomaculatus | 165–183 | 53–63 | no | absent but a small spot present | white | yellow bands extending onto ventral scale edges |
| jara | 167–188 | 52–74 | no | absent | whitish-grey | overall speckled with small yellow spots |
| laoensis | 163–187 | 60–76 | yes | present /yes | whitish | yellow-white bands extending onto ventral scale edges |
| travancoricus | 176–206 | 64–76 | yes | present /scarcely | greyish, mottled with brown | yellow bands extending onto ventral scale edges |
| striatus | 154–195 | 35–50 | no | present / yes | whitish | whitish-brown or yellowish bands extending onto ventral scale edges |
| zawi | 179–207 | 45–75 | yes | absent | whitish-brown | short white bands not extending onto ventral scale edges |
In a recent study on population systematics of the L. aulicus complex
Our comparison of the original information of fasciolatus Shaw, with data from the two morphotypes identified by Ganesh and Vogel (l.c.), showed the closest match with L. anamallensis (see Table
Based on these consistencies of its morphological characters, colour and pattern, we consider Coluber fasciolatus Shaw, 1802 and Coluber hebe Daudin, 1803 as conspecific with Lycodon anamallensis Günther, 1864, including its synonym Lycodon osmanhilli Taylor, 1950.
With the reallocation of the taxon fasciolatus Shaw to the genus Lycodon we follow the lead of H.
Description
According to
Distribution
Lycodon fasciolatus seems to be widespread in southern peninsular India and recent records from the Coromandel region support the possible demarcation of the type locality in this area. According to
Some old records mention a specimen under the name Coluber or Coryphodon fasciolatus (
Recently,
Remarks
It is undisputed that Blyth (l.c.) considered the resemblance of C. vittacaudatus to C. fasciolatus in the sense of Shaw, so he clearly did not mean what is now commonly referred to as the Banded racer. Because of our clarification that C. fasciolatus is a wolfsnake, all previous assumptions concerning the identity of Coluber vittacaudatus Blyth are questionable.
Platyceps plinii
Coluber. –
Coluber pictus
Daudin, 1803, p. 347–349, “La Coleuvre Peinte”, type locality “Casem-Cottah, au Bengale”, based on
[Natrix] Plinii Merrem, 1820, p. 101 f., “halbbandierte [Natter]”, type locality “Bengalen”, restricted here to “Casemcottah” (= Kasimkota village, Andhra Pradesh state, India; see Appendix Note 1), based on
Coluber plinii. –
Coluber curvirostris
Cantor, 1839: 51, type locality “Bengal”, holotype not located, an unpublished original sketch depicting the iconotype is kept in the Bodleian Library (Oxford, U.K.), [reproduced here in Fig.
Coluber fasciolatus. –
Coronella baliodeira. –
Coryphodon fasciolatus. –
Tyria
[fasciolata]. –
Zamenis fasciolatus. –
A.
[rgyrogena] rostrata Werner, 1924, p. 51, fig, 4a–c, type locality “Argentinien” [in error], holotype NMW 18160;
Argyrogena fasciolata. –
Argyrogena fasciolatus
[sic]. –
Platyceps plinii. – this work.
The preceding list of synonyms and chresonyms contains quotations which to our best knowledge refer to the snake species so far known as Banded racer (Argyrogena fasciolata auctt.) in the sense of
Coluber curvirostris was based on a single specimen from “Bengal” described by Theodore


Extract from a previously unpublished original water-colour painting from Brian H. Hodgson’s collection of drawings kept in the Zoological Library of the Natural History Museum, London, U.K. The depicted specimen from the „Central hilly region “of Nepal, determined by

Platyceps plinii comb. nov. Previously unpublished original painting (no. 7) of the holotype of Coluber curvirostris Cantor, 1839 from Cantor’s original manuscript entitled “Indian Serpents–Innocuous–Collected, figured & described (1831–1837)”, kept in the Bodleian Library, Oxford, U.K. (see Appendix, Note 2). Reproduced with permission from the Bodleian Library, Oxford, U.K.
Franz
We also tracked down two older and previously overlooked or wrongly assigned names that clearly refer to what is currently understood to be the Banded racer, viz. Coluber pictus Daudin, 1803 and Natrix plinii Merrem, 1820. Again both names are entirely based on the same description in
From the mid of the 19th century onwards both names fell into oblivion as potential synonyms of the Banded racer. This was probably also due to
Coluber pictus Daudin, 1803 however, is threatened for reasons of homonymy by an older name, Coluber pictus Gmelin, 1789, the accepted original combination for the Painted bronzeback (now Dendrelaphis pictus, see ICZN 1958). Therefore, Coluber pictus Daudin, 1803 represents a younger primary homonym of Coluber pictus Gmelin, 1789 and is unavailable (ICZN 1999, Art. 57.2; see also above the list of synonyms and chresonyms for further homonymous names).
To our best knowledge, Natrix plinii Merrem, 1820 is the oldest available name for the snake species which is commonly now regarded as Banded racer. Merrem (l.c.) simplified the type locality to “Bengal”, but according to
Type material
We could not locate the two type specimens of Natrix plinii Merrem, 1820 among Russell’s dry and wet preserved material stored in the collections of the Natural History Museum, London, U.K. A wet preserved topotypic male (
The same applies to the type specimen of Coluber curvirostris Cantor, 1839 from “Bengal” which is not yet located. Although some of Cantor’s original specimens are known to be stored in the Natural History Museum, London (
Description
Based on our own data and summarized information from literature (
Variation
Pholidosis: Head 2.17–2.14 times longer than broad (males 1.17–2.14, females 1.22–2.05), canthus rostralis moderate developed. Rostral nearly twice as broad as high, clearly visible from above. Internasals usually shorter than prefrontals, sometimes equal in length in, e.g.
Predominantly eight supralabials, last two or three being larger (longer), fifth highest; nine in
Dorsal scale reduction formula summarized from 31 examined specimens (see Appendix
2+3, 3+4, or 10+11 (6–11) 2+3, 3+4, 7+8, 8+9, 9+10, or 10+11 (65–131) [79–133]
25 -------------------------------- 23 -------------------------------------------------------------------
3+4, or 10+11 (5–9) 2+3, 3+4, 7+8, 8+9, 9+10, or 10+11 (63–131) [82–129]
2+3, 3+4, 4+5, 8+9, 9+10, or 10+11 (68–145) [105–137]
21 ------------------------------------------------------------------------
1+2, 2+3, 3+4, 4+5, 7+8, 8+9, or 9+10 (68–142) [107–138]
4+5, 6+7, 7+8, 8+9, or 9+10 (111–192) [157–174] 7+8 (188–194)
19 ------------------------------------------------------------- 17 --------------------------- 15.
3+4, 4+5, 7+8, 8+9, or 9+10 (112–191) [156–178] 7+8, or 8+9 (190–195)
Six examined specimens (
Hemipenis (based on ZSI-CZRC-V-6416, Pune, left organ, Fig. 11 )
The hemipenis reaches up to 13th subcaudals and is 36.9 mm in length and 8.2 mm maximum width at midbody and apex. It is divided into three distinct areas; the proximal area (nearly 30% of the total hemipenial length) is smooth without any ornamentation, middle zone (about 25% of the total hemipenial length) is with evenly scattered spinules and the apical calyculated portion (45% of the total hemipenial length). No enlarged spines are present, but the size of spinules at the proximal end are slightly larger than the ones at the distal end. The calyculated area can be further divided almost equally into proximal half with large calyces and distal half with smaller and denser calyces. The sulcus spermaticus is single, bounded with thick walls, runs straight across its length and opens into a delta (3 mm wide at apex, 1.9 mm in length), which is nude. The delta is subapical in position and the apex is calyculated, edged with fewer papillae. At the sulcate side, the calyces are larger proximally and gradually smaller and denser towards the distal end. The distal calyces are scalloped and edged with papillae. Along the asulcate side the spinous area starts more distally than the sulcate and lateral side. After the spinous zone, there are 2–3 rows of large calyces followed by densely packed calyces towards the apex. The proximal large calyces (9–10 in number) are of uneven size (mostly rounded or pentagonal) and among them the distal ones are scalloped. The large scalloped calyces are of 2.2–2.7 mm in length and 1.9–2.5 mm in width.
Dentition (based on ZMB 8053, male, Bengalen)
15/15 maxillary teeth, the anterior 13/13 precranterian teeth are increasing in size posteriorly and are followed by a small diastema and 2/2 enlarged roundish cranterian teeth without groove. All maxillary teeth are slightly curved posteriorly, without significant interspaces. Medial to the precranterian teeth are single replacement tooth at different growth stages. One to two replacement teeth per cranterian tooth are found posteromedially to each tooth, showing different growth stages. 11/11 palatine teeth, decreasing in size posteriorly. All are curved posteriorly shortly above the base. Lateral to the palatine teeth there is a single replacement tooth at different growth stages. No significant interspace exists between the different palatine teeth. Posteromedial process of palatine large, expanding two pterygoid teeth. 13/12 pterygoid teeth, decreasing in size posteriorly. All are curved posteriorly shortly above the base. Lateral to the pterygoid teeth there is a single replacement tooth at different growth stages. No significant interspace exists between the different palatine teeth. The posterior 57% of the pterygoid are without teeth. 17/17 mandibular teeth, increasing in size up to tooth 5/5 and from there decreasing in size posteriorly. All are slightly curved posteriorly. Medial to the mandibular teeth there are 1–2 replacement teeth at different growth stages.
Variation in dentition
Four specimens from Pakistan (
Dimensions and proportions
The body is robust, moderately stout, roundish in cross-section at midbody. Head moderately pointed, barely distinct from neck. Eye large, 16–24% of head length. Male specimens grow slightly larger than females; the longest examined specimens are from Sind, Pakistan including a male (
Colour and pattern (Fig. 9 A–K and Fig. 10 A–J )
Dorsal colour and pattern show a significant ontogenetic change. In juvenile and subadult specimens the dorsal ground colour is olive-brown to light brown, the head dorsally slightly darker with narrow elongated irregular shaped whitish markings. Along the body up to 70 narrow whitish/black or dark brown stippled crossbars, usually two scales wide and in contact with lateral edges of ventrals. The first band is separated from the posterior border of parietals, the interspace between bands is three to four dorsal scales wide. The intensity of bands decreases posteriorly and the contours become blurred near to the vent. Dorsocaudally the bands are dissolved in an irregular fine pattern of light and dark points. Temporals, preoculars and supralabials can show small whitish spots. From about 600 mm total length the colour and pattern of young specimens begin to fade and disappear gradually. Adult specimens are dorsally dark brown to dark reddish-brown without any markings on head, body or tail. The ventral side of head, body and tail in all age classes is patternless yellowish cream.
Distribution
Platyceps plinii is commonly believed to occur from Pakistan in the West to Bangladesh in the East with a North to South distribution from Nepal to Sri Lanka (see e.g.
According to our current knowledge, Platyceps plinii is only known with certainty from India and Pakistan. In India it is recorded from the states of Andhra Pradesh (Kasimkota [Casemcottah, type locality], Eluru, Nallamala Hills, Pulicat Lake, Rajahmundry, Rollapenta, Sundipentha/Sikharam, Thummalapalle mine and Visakhapatnam), Bihar (Munger, Patna, Tinpahar and Rajmahal), Chhattisgarh (Pharsabhar), Goa (Bondla Wildlife Sanctuary and Panaji), Gujarat (Baroda, Bharuch, Bhavnagar, Dahod, Dangs, Gandhinagar, Girnar Hill, Gir Forest, Kala Gadba, nr. Mahal, Mehsana, Panchmahal, Sabarkantha, Samot, Surat, Valsad, Vansda National Park and Vijapur), Haryana (Ambala), Jharkhand (Hazaribag District), Karnataka (Balekola, Chincholi Forest, Belgaum, Coimbatore, Coorg, Hosar, Mysuru and Shivamogga), Kerala (Muthanga in Wayanad Wildlife Sanctuary), Madhya Pradesh (Bhopal district, Barkatullah University Campus, Dewas, Dumna Nature Park, Gwalior, Indore, Jabalpur, Kanha National Park, Katra Hills, Mandla, Pachmarhi Biosphere Reserve, Sagar, Satpura Tiger Reserve, Shahdol, Sita Hill and Ujjain), Maharashtra (Akola district, Ahmednagar, Ale nr. Narayangon, Atpadi, Aurangabad, Bassein Fort, Bohali, Borivali (Gorai) mangroves, Buldhana district, Chink Hill, Dahanu Forest Division, Deolali, Devagad, Ghorawadi nr. Talegaon, Jalgaon, Jawhar, Juvem, Kaas Plateau, Khandala, Kolhapur district, Konkan region, Kurduvadi, Malegaon, Marol, Mokhada, Mumbai, Nagar, Nagpur, Nanded, Navegaon National Park, Navi, Panchgani, Parel, Parvati-Pachgaon Hills, Powai, Pune, Ranidoh region, Thana, Vidarbha region, Vidyanagari, Visapur and Yavatmal), Odisha (Gandhamardan Hills, Nandankanan, Choudwar, Similipal Biosphere Reserve, Baripada and Bhadrak), Puducherry, Rajasthan (near Hemavas Dam, Pratapgarh district and Ramri), Tamil Nadu (Chennai and Hosur Hills), Telangana (Farahabad, Hyderabad region, Ippalapally, Jannaram, Kerimeri, Khanapur, Kinnerasani, Old Bowenpally, Rushulacheruvu, Tarnaka and Vijayapuri), Uttar Pradesh (Mathura region, Faizabad, Prayagraj and Varanasi), West Bengal (Dum Dum, Durgapur, Kolkata and Panchet Hill). The westernmost published record of the Banded racer comes from the Girnat Hill or Girnagar in the Junagadh district of Gujarat, approximately 70°31’ East (

Platyceps plinii comb. nov. Previously unpublished original colour-painting from the Thomas Hardwicke collection of sketches (Volume II, sketch 63) kept in the Zoological Library of the Natural History Museum, London, U.K. (see also

Platyceps plinii comb. nov. in life from various parts of India. Adults: A. ZSI-CZRC-6384 (male, SVL: 491) from Baripada, Odisha state, B. Uncollected specimen from Pune, Maharashtra state, C. ZSI-CZRC-V-6416 (male, SVL: 815) from Pune, Maharashtra state, and D. Uncollected female from Amaravati, Maharashtra state. Hatchlings/juveniles: E. ZSI-CZRC-V-6329 (subadult male, SVL: 355) from Choudwar, Odisha state, F. Uncollected specimen from Pune, Maharashtra state, G. Uncollected specimen from Amboli, Maharashtra state, and H. Uncollected specimen from Yavatmal, Maharashtra state. SVL in mm.

Platyceps plinii comb nov. Variation of neck pattern in specimens from various parts of India. Adults: A. NCBS-AQ492 (female, SVL: 820) from Bangalore, Karnataka state B.

In Pakistan confirmed records are known from the Indus plain in Sindh province (nr. Badin, Hala, Jati, Lakarna, Makli Hills, Mohenjo-daro, Pir Patho, Raj Malk, Sonda, and Tatta) and some authors mentioned southern and central Punjab without specific locality (e.g.
Unconfirmed or erroneous distribution records
Sporadically the Banded racer is mentioned for the Himalayan region in northwestern and northeastern India.
Based on water-colour sketch no. 61 from the Hardwicke collection,
When it comes to the distribution of the Banded racer in Sri Lanka, there is a consistent reference to
In several publications, especially in overview works and faunal lists, the Banded racer is mentioned for Bangladesh. This is probably because in older publications (e.g.
From Nepal, the Banded racer was first mentioned by
Based on records of the Banded racer in Bengal,
Habitat and natural history
Platyceps plinii is widespread over the Indian Subcontinent (see above, Map 1 and Appendix


The vertical distribution ranges from sea level to over 1000 m altitudes, with about 85% of all finds below 600 m a.s.l. A few unusually high sites are known from the states of Madhya Pradesh (Pachmarhi Biosphere Reserve, 1053 m a.s.l.) and Maharashtra (Kaas Plateau, 1191 m a.s.l., and Panchgani, 1258 m a.s.l.) (see Appendix
It is a mainly terrestrial species but also observed to be a good climber (
It is a diurnal species that is only exceptionally observed active during the night and known for its defensive and pugnacious behaviour. If threatened or captured it erects itself, flattens the forebody and furiously attacks when further provoked which one can confuse with a cobra (
The Banded racer is an active hunter that usually kills its prey by constriction or body pressure. Its prey consists of small mammals like rats, mice, shrews and bats, but also frogs, lizards, birds, worms, and for juveniles insects are part of the food spectrum (
Mating of the Banded racer was observed between late winter and mid-monsoon (
Manhas (2015a) observed the Banded racer as roadkill during monsoon in Bhopal district of Madhya Pradesh state and
For Pakistani specimens,
Platyceps josephi
“The Racer”. –
Zamenis fasciolatus. –
Coryphodon fasciolatus. –
Argyrogena fasciolatus
[sic]. –
Argyrogena fasciolata. –
Coluber fasciolatus. –
Platyceps josephi sp. nov. – this work.
Diagnosis
A medium sized (maximum total length 951 mm) snake with countersunk lower jaw; dark brownish dorsum; head with irregular white spots, two slanting roughly “∏” shaped white markings with black edges on either side, starting on the back of the head (behind parietals) extending into the body, almost the length of head; 13–18 prominent white bands in the anterior region of the dorsum in both juveniles and adults; 34–48 total bands on the body in both juveniles and adults; 23:21(exceptionally 23):16–18 smooth dorsal scale rows; 189–218 ventrals (males: 192–197; females: 189–216); 76–88 subcaudals (males: 83–88; females: 76–88); cloacal plate divided; tail without bands and underside creamish. Its dentition is characterized by 12–16 maxillary teeth, the last two enlarged and separated by a diastema, 9–11 palatine teeth, 14–15 pterygoid teeth and 15–16 mandibular teeth.
Platyceps josephi sp. nov. shows most similarities with its sister taxon P. plinii in regard to pholidosis and colour pattern but can be distinguished from the latter by its lower number of midbody dorsal scale rows (21 vs. 23), its lower mean value of ventral scales (202 vs. 214), its lower mean value of subcaudal scales (82 vs. 87) and its lower mean of the sum of ventral and subcaudal scales (285 vs. 304), the presence of the clearly demarcated two slanting roughly “∏” shaped white markings on the back of head vs. absence of such markings and distinct white bands on the dorsum in both juveniles and adults vs. ontogenetic change, i.e. bands present in juveniles only, but usually absent or faded in adults. Additionally, Platyceps josephi sp. nov. differs from P. plinii in variation of mitochondrial DNA sequences. With pairwise uncorrected p-distances varying 4–5% in cytb & ND4 and 3% in 16S. It is also clear from our thorough verification of distribution that these two species only have a minor range overlap in northeastern Tamil Nadu (Fig.
Holotype
NCBS AU-732, adult female, from Tuticorin, Tamil Nadu state, India (8.75442° N, 78.18482° E, 5 m a.s.l) collected by Naveen Joseph on 26th February 2017 (Figs



Platyceps josephi sp. nov. in life from various parts of Tamil Nadu state, India: A. Holotype NCBS-AU732 (female, SVL: 757) from Tuticorin, B. Paratype NCBS-AU733 (female, SVL: 608) from Tuticorin, C. Paratype
Paratypes
Referred specimens
ZSI-CZRC-6521, adult female, from Tuticorin, Tamil Nadu state, India (8.73448° N, 77.97889° E, 33 m a.s.l.) collected by Naveen Joseph on 12th June 2017; ZSI-CZRC-6522, adult female, from Vilathikulam, Tamil Nadu state, India (9.125623° N, 78.176763° E, 20 m a.s.l.), collected by Ahmed Jerith and Naveen Joseph on 2nd February 2018; ZSI-CZRC-7358, sex unknown, from Anaikatti, Coimbatore district, Tamil Nadu state, India (11.11221° N, 76.75795° E, 581 m a.s.l.), collected by Jins VJ on 23rd Octobe
Etymology
The specific epithet is a patronym of late Mr Naveen Joseph. Naveen was a naturalist from Tuticorin, well known for his research on reptiles, particularly snakes in that region. He was a friend of VD, SN, and GM and helped them collect specimens of the new species. Suggested English name: Joseph’s racer. Vernacular name: In various parts of Tamil Nadu state this snake is called by the name “Odugali Pambu” “ஓடுகாலிப்பாம்பு”. It is a portmanteau word in Tamil language “Odugali” is often used to address someone “who doesn’t stay at home and elopes” and “Pambu” is the word for “snake”.
Description of the holotype
Morphometric and merestic data are provided in Table
Meristic and morphometric (in mm) data for examined specimens of P. josephi sp. nov. Symmetrical head characters are given in left/right order.
| Inventory no. | NCBS AU-732 holotype | ZSI-CZRC-6639 paratype |
|
NCBS AU-733 paratype | NMW 25465:2 paratype | NMW 25465:1 | ZSI-CZRC-6522 | ZSI-CZRC-6521 | ZSI-CZRC-7358 |
| Sex | female | male | female | female | male | female | female | female | unknown |
| SVL | 757.6 | 574.3 | 592.0 | 608.5 | 440.0 | 255.0 | 574.5 | 722.2 | NA |
| TaL | 175.5 | 167.1 | 162.2 | 89.9 | 142.0 | 60.0 | 170.5 | 228.9 | NA |
| TL | 933.1 | 741.5 | 754.2 | 698.4 | 582.0 | 315.0 | 745.0 | 951.1 | NA |
| HW | 12.4 | 11.9 | 10.8 | 13.5 | 11.6 | 7.9 | 13.4 | NA | NA |
| HL | 24.1 | 20.0 | 21.6 | 22.7 | 14.5 | 10.7 | 23.0 | NA | NA |
| ED | 4.1 | 3.4 | 3.9 | 3.8 | 3.3 | 2.7 | 3.7 | NA | NA |
| RW | 4.8 | 4.8 | 4.2 | 4.6 | 3.8 | 2.4 | 4.1 | NA | NA |
| FL | 6.4 | 6.1 | 5.6 | 5.3 | 5.2 | 4.3 | 5.9 | NA | NA |
| FW | 4.7 | 3.8 | 4.3 | 3.7 | 3.7 | 3.0 | 4.2 | NA | NA |
| PrefL | 2.0 | 2.3 | 2.2 | 2.2 | 2.2 | 1.6 | 2.0 | NA | NA |
| InL | 1.2 | 1.6 | 1.6 | 1.5 | 2.1 | 1.5 | 1.9 | NA | NA |
| E-S | 8.2 | 7.6 | 7.2 | 7.7 | 6.3 | 4.6 | 7.3 | NA | NA |
| ParL | 6.3 | 5.7 | 6.0 | 6.0 | 5.7 | 4.5 | 5.8 | NA | NA |
| ParW | 4.5 | 4.2 | 4.2 | 4.1 | 3.9 | 3.0 | 4.6 | NA | NA |
| DSR | 23/21/18 | 24/23/17 | 23/21/17 | 23/21/17 | 21/21/17 | 23/23/17 | NA | 23/21/16 | NA |
| PreVen | 3 | 1 | 2 | 0 | 2 | 1 | 3 | 2 | NA |
| Ven | 203 | 192 | 195 | 208 | 197 | 216 | 204 | 189 | 218 |
| Subc | 76 | 82 | 77 | 84 | 88 | 79 | damaged | 88 | NA |
| TS | 283 | 276 | 275 | 293 | 288 | 297 | NA | 280 | NA |
| PrO | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 |
| PoO | 2/2 | 2/2 | 2/2 | 2/2 | 2/3 | 2/2 | 2/2 | 2/2 | 2/2 |
| T | 2+2/2+3 | 2+3+4/2+2+4 | 2+3/2+3 | 2+3/2+3 | 2+2+3/2+3+4 | 2+3+4/2+2+4 | 2+3+4/2+2+4 | 2+3/damaged | 2+3/2+3 |
| SupL | 8/8 | 8/8 | 8/8 | 8/8 | 8/8 | 8/8 | 8/8 | 8/8 | 8/8 |
| SubL | 9/9 | 10/10 | 10/10 | 10/10 | 11/11 | 10/10 | 10/10 | damaged | 10/10 |
Body subcylindrical, dorsoventrally flattened. Head ovate, barely wider than the anterior end of body. Total lenghth 934 mm; snout-vent length 758 mm; tail length 176 mm; ratio tail length/total length 0.19; head length 24.0 mm; rostral large, 2.2 times broader than high, not protruding, rounded in dorsal view, and wedged in between internasals; the latter smaller (1.5 mm) and distinctly shorter along median suture than prefrontals (2.5 mm); mid-line suture between internasals and pre-frontals in straight line; distance from posterior tip of rostal to anterior edge of frontal 4.2 mm, the latter bell-shaped, 6.4 mm long with a maximum width of 4.7 mm; interocular width 7.0 mm; parietals 6.3 mm long, outer lateral and posterior margins of parietals surrounded by 13 scales between the upper postoculars; nasal completely divided, in contact with first and anterior half of second supralabial; nostril large in anterior shield; distance between posterior margin of nostril and anterior border eye 4.2 mm; loreal slightly longer (1.5 mm) than high (1.4 mm), in contact with posterior part of second and anterior half of third supralabial; preocular single, reaching top of head, touching frontal; presubocular squarish, smaller than preocular and loreal, in contact with third and fourth supralabial; 8/8 supralabials, fourth and fifth entering the eye, fifth highest; 2/2 postoculars, upper slightly broader, lower higher, lower postocular in contact with fifth supralabial; temporals in 2+3/2+3 rows, lower anterior slightly larger than upper; 9/9 sublabials, sixth largest, first five in contact with anterior inframaxillaries on both sides; anterior pair of inframaxillars broader and slightly longer (6.1 mm) than posterior pair (5.6 mm), which is distinctly separated by five inserted gulars; five gulars in a row between posterior edge of anterior inframaxillars and preventral; three preventrals and 203 ventrals; anal scute divided, right overlapping left; 76 paired subcaudals; terminal spine blunt; dorsal scales smooth, arranged in 23/21/17; apical pits present, but not consistently on all the dorsal scales, single or paired on dorsal part from the neck to near sacral region, single or triple pits present on dorsocaudal scales.

Platyceps josephi sp. nov., dorsal view of preserved paratypes from Tamil Nadu state, India: A. ZSI-CZRC-6639 (male, SVL: 574), from Karur, B. NCBS-AU733 (female, SVL: 608) from Tuticorin, C. NMW 25465:2 (male, SVL: 440) from Salem, and D.

Platyceps josephi sp. nov. Variation of neck pattern in specimens from various parts of Tamil Nadu state, India. Adults: A. Paratype, ZSI-CZRC-6639 (male, SVL: 756) from Karur, B. Paratype,
Dorsal scale reduction formula:
-3(6) +10(64) -10(67) +10(82) 10+11(87) 3+4(133) 8+9(151) –
23-------21-----------23---------21-----------23--------------21------------19------------17----------18(203).
-4(8) +11(65) -11(69) +11(84) -11(88) 3+4 (133) 8+9(150) +2(202)
Dentition (see Fig. 20 )
14/16 maxillary teeth, the anterior 12/14 precranterian teeth are increasing in size posteriorly (from 0.9 to 1.6 mm length) and are followed by a diastema which is as long as the socket of the last precranterian tooth, and 2/2 slightly enlarged roundish cranterian teeth (~1.9 mm long) without groove. All maxillary teeth are slightly curved posteriorly, without significant interspaces. Medial to the precranterian teeth are single replacement tooth at different growth stages. One to 2 replacement teeth per cranterian tooth are found posteromedially to each tooth, showing different growth stages.
9/9 palatine teeth, decreasing in size posteriorly. All are curved posteriorly shortly above the base. Lateral to the palatine teeth there are single replacement tooth at different growth stages. No significant interspace exists between the different palatine teeth. Posteromedial process of palatine large, expanding two pterygoid teeth.
14/14 pterygoid teeth, decreasing in size posteriorly. All are curved posteriorly shortly above the base. Lateral to the pterygoid teeth there are single replacement tooth at different growth stages. No significant interspace exists between the different palatine teeth. The posterior 56% of the pterygoid are without teeth.
15/15 mandibular teeth, increasing in size up to tooth 6/6 and from there decreasing in size posteriorly. All are slightly curved posteriorly. Medial to the mandibular teeth there are 1–2 replacement teeth at different growth stages.
Hemipenis (based on ZSI-CZRC-6639, paratype, Karur, right organ, Fig. 19)
The hemipenis reaches up to 12th subcaudals and is 24.3 mm in length with a maximum width of 5.8 mm at 1/3rd of the proximal end. It is divided into three distinct areas; the proximal area (nearly 1/4th of the total hemipenial length) is smooth without any ornamentation, middle zone (about 28% of the total hemipenial length) is with spinulae and the apical calyculated portion (48% of the total hemipenial length). The spicules at the midbody of the hemipenis gradually reduce in size from proximal to the distal end. The calyculated area can be further divided almost equally into proximal half with large calyces and distal half with smaller and denser calyces. The sulcus spermaticus is single and runs straight across its length and opens into a wide and elongate naked area (3.3 mm) extending laterally from the proximal to distal end extending to the apex. The opening of the sulcus forms a triangular area and ends apically. The sulcus is bounded with thick walls. The calyces are larger proximally and gradually smaller and denser towards the distal end. The calyces distally are edged with papillae. Along the asulcate side, the spinous area starts more distally than the sulcate and lateral side. The proximal calyces are wider and elongated, with eight to nine well defined calyces extending towards the lateral side and the distal part of these calyces are scalloped. The calyces towards the distal part are with more of papiliated edges and are similar in micro-ornamentation with the sulcate side.


Variation among the paratypes
See Table
NCBS-AU733 differs from the holotype in having a slightly darker frontal and suprocular scales, presence of irregular white spots in the temporal scales, and a small white line along the midline between the two prefrontals. The first band on the body is separated from the “∏” shaped markings by 2 scales on both the sides along the lateral side. 46–48 bands on the dorsum from neck to vent, anterior 18 bands are prominent with white and black edges which fades and become wider towards the vent.
Variation in dentition
The male paratype from “Salem” (NMW 25465: 2) show 12/12 maxillary teeth followed by a distinct diastema which is 50% longer as the socket of the last precranterian tooth and two enlarged cranterian teeth, 11/11 palatine teeth with a posteriomedial process of palatine expanding three pterygoid teeth, 15/14 pterygoid teeth with 41% of the posterior pterygoid without teeth, and 15–16 mandibular teeth, increasing in size up to tooth 7/7 and from there decreasing in size posteriorly.
Pholidosis
Head 1.25–1.99 times longer than broad (male 1.25–1.67, females 1.35–1.99). Rostral twice as broad as high and visible from above. Internasals (in midline suture) usually smaller than prefrontals. Frontals 1.32–1.59 times longer than the maximum width (male 1.40–1.59, females 1.32–1.42). Posterior border of parietals uneven in shape at the median suture forming an acute angle in ZSI-CZRC-6639 and NCBS-AU733 or both parietals uneven forming a slightly acute angle in
Dorsal scale rows (DSR), usually arranged in 23–24/21, rarely 23/17 or sometimes 18 rows along the trunk. One male specimen from Karur shows anterior 24 DSR; Midbody DSR predominantly 21 except in one specimen ZSI-CZRC-6639 with 23 DSR and two specimens NCBS-AU732 and NCBS-AU733 shows 18 DSR and one specimen ZSI-CZRC-6521 shows 16 DSR in posterior part of the body.
Dorsal scales with single or paired apical pits. Supracaudal scales usually with one to three pits on the first three to five scales followed the supracaudal reduction. In male (ZSI-CZRC-6639), consistently two pits from the midbody to the last supracaudal scale.
Dorsal scale reduction formula summarized from five examined specimens (see Appendix
4+5(6) 3+4(6–13) 2+3 or 3+4(116–133) 7+8 or 8+9 (144–151)
24 -------- 23 --------------------- 21-------------------------- 19 --------------------------- 17.
– 2+3 or 3+4(6–13) 3+4 (115–133) 7+8 or 8+9 (142–151)
Two examined specimens (NCBS-AU732, holotype, and NCBS-AU733) and one specimen (ZSI-CZRC-6521) show an additional lateral increase to 18 and reduction to 16 dorsal scales involving rows 1–3 between 95 and 99% of ventral scales respectively.
Dimensions and proportions
The body robust, moderately stout, roundish in cross section at midbody. Head moderately pointed, barely distinct from neck or sometimes slightly distinct. Eye large with black round pupil, 16–25% of head length. The longest examined specimens are from Karur (ZSI-CZRC-6639), a male specimen with 740 mm and a female from Tuticorin (ZSI-CZRC-6521) with 951 mm total length. The smallest specimens are from Salem including a male (NMW 25465: 2) with 582 mm and female (NMW 25465:1) with 315 mm total length. The tail/body length ratio ranges from 0.15 to 0.32 (males 0.29–0.32, females 0.15–0.32).
Distribution
Platyceps josephi sp. nov. is so far only known from Tamil Nadu state, India (see Fig.
Habitat and natural history
Platyceps josephi sp. nov. mostly inhabits open habitats with sandy or rocky patches in grasslands and scrublands in both inland and coastal areas of Tamil Nadu from elevations between 10 and 580 m a.s.l. (see Fig.

Platyceps josephi sp. nov. is a diurnal snake, terrestrial and swift in locomotion. However, there is a report of arboreal behaviour under artificial conditions (
The diet of the P. josephi sp. nov. chiefly consists of geckos, lizards and small rodents. It is observed to kill the prey by constricting and/or crushing the prey against ground or tree trunk. In all captive observations, it swallows the prey from the head first. It is an oviparous snake with few records of clutch sizes, 7–12 eggs (
Platyceps josephi sp. nov. is reported only from one protected area (Megamalai) in its known range. The species faces a number of threats across its distributional range, including habitat destruction, because the grasslands in southern Tamil Nadu are being actively converted into plantations, farmlands and urbanisation. Although there are records from such converted plantations and human settlements,the species’ ability to adapt and its reproductive success is not known. Rocky habitats in Madurai region are also highly affected by the mining activities and road traffic is another important threat to P. josephi sp. nov. Three out of the seven specimens collected in this study were roadkilled and six other uncollected roadkilled specimens were observed from various parts of Tamil Nadu between 2017 and 2020. The Area Of Occupancy (AOO) of P. josephi sp. nov. is 72.000 km2 and Extent Of Occupancy (EOO) is 70,698 km2. Even though this is a relatively large area of distribution for a species, our field surveys and records suggest that this species has patchy distribution within its range. Also, much of the habitat in these regions where P. josephi sp. nov. is reported are under severe threats like conversion of grasslands to farmlands, widescale monoculture plantations (Eucalyptus sp.) and urbanization. Given this information, we suggest that P. josephi sp. nov. should be considered a species in the Vulnerable category according to the IUCN criteria.
Although local envenoming by congeners is reported for P. rhodorachis and P. najadum (
Juvenile Platyceps josephi sp. nov. can easily be confused with juveniles of Platyceps plinii but may be differentiated by the two slanting roughly “∏” shaped markings on the back of the head. Platyceps josephi sp. nov. is found in sympatry with the Common cobra (Naja naja) and the Common ratsnake, Ptyas mucosa, and can potentially also be confused with these two species.
Differences between Platyceps josephi sp. nov. and South Asian congeners
Platyceps josephi sp. nov. is distinguished clearly from P. bholanathi, P. mintonorum, P. noeli, P. rhodorachis, P. sindhensis and P. ventromaculatus by its higher number of midbody dorsal scale rows (21 vs. 19) and from P. gracilis by its lower mean value of ventral scales (202 vs. 214), its lower value of subcaudal scales without overlapping (88 maximum vs. 118 minimum), its different neck pattern (whitish ∏-shaped marking vs. a yellowish-cream and black edged V-shaped marking), and by differing dorsal body pattern (irregular whitish transverse bars stippled with black vs. dorsal yellowish-cream and black-edged transverse bars). Platyceps josephi sp. nov. shows similarities with P. plinii with regard to pholidosis and colour pattern but can be distinguished from the latter by its lower number of midbody dorsal scale rows (21 vs. 23), its lower mean value of ventral scales (202 vs. 214), its lower mean value of subcaudal scales (82 vs. 87), and its lower mean of the sum of ventral and subcaudal scales (285 vs. 304).
Comparision of Osteology
The first detailed description of osteological features of Platyceps plinii were based on a male and a female specimen (
In his comparison,
Some of the osteological characters used by
Recently,
In contrast to the former studies on Argyrogena mentioned above, material examined by us lead to different results (see also Appendix
We found differences within the distal ends of the lateral processes of the premaxilla, which either are tapering into a single tip in P. plinii comb. nov., P. josephi sp. nov., and P. rhodorachis, expanded and divided into two tips in P. florulentus or expanded into a stout end in P. rhodorachis as well as in P. ventromaculatus. Because of this considerable intraspecific variation, we regard the form of the lateral processes of the premaxilla as an inappropriate character to delimit Argyrogena from Platyceps.
Furthermore,
Wilson (l. c.) mentioned that in Argyrogena the nasal shield posteriorly expands dorsolaterally to cover the nasal capsule more completely as in the other Coluber (sensu lato). Our measurements show that the nasal shield of the Banded racer tends to be shorter (TLln/Lns range 40%–50%) than in other examined Platyceps spp. (TLln/Lns range 56%–77%) and therefore covers only a smaller part of the nasal capsule. Our comparison of the nasal shields revealed that it covers in P. plinii comb. nov. the posterior part of the nasal capsule completely, whereas in the other examined Platyceps spp. (excluding josephi) only the middle part of the nasal capsule is enclosed. This resulted in a larger gap between the nasal shield and the frontal. Platyceps josephi sp. nov. shows an intermediate state regarding this character as the nasal shield do enclose the posterior part of the nasal capsule but shows a larger gap than observed in P. plinii comb. nov. This character seems to be highly influenced by the lifestyle of the species and varies considerably interspecifically (
Another character listed by Wilson (l. c.) as typical for Argyrogena deals with the pterygoid teeth row, which converges posteriorly in the Banded racer but diverges in Coluber (sensu lato). We cannot follow this observation as in all examined specimens the teeth row follows the medial edge of the pterygoid bone and therefore always converge posteriorly.
Wilson (l. c.) also described the pterygoid processes at the basioccipital which he mentioned as clearly visible in e.g. Platyceps karelini, P. ventromaculatus, Dolichophis jugularis and Hierophis viridiflavus but lacking in Argyrogena. In Platyceps spp. examined by us, we found a high variability of this character, ranging from a basioccipital without structures, with three or five small tips or sometimes with a high crest. Our comparison of different development stages of D. jugularis, H. gemonensis and H. viridiflavus show that in juveniles such structures are lacking while in adults, high crests and tips appear and the expression of this character is very likely age-dependent.
Differences in midbody vertebrae morphology of racer species is highlighted as a character of phylogenetic significance (
According to
In summary, it is evident that Argyrogena shares most osteological characters with Platyceps. Previous use of skeletal traits to underpin the distinctness of the genus Argyrogena is shown invalid, with the exception of the form of the nasal shield as the only distinguishing character to the previous known Platyceps spp. But with respect to the latter character, Platyceps josephi sp. nov. show an intermediate state and linked the Banded racer with Platyceps. We therefore consider Argyrogena Werner, 1924 a junior subjective synonym of Platyceps Blyth, 1860 and relegate the Banded racer (auctt.) to the genus Platyceps in the new combination Platyceps plinii (Merrem, 1820) (see the also conclusions in previous chapters).
Identification key to South Asian Platyceps (modified from Schätti et al. 2014)
| 1a. | Midbody dorsal scales arranged in 23 rows | plinii |
| 1b. | Midbody dorsal scales arranged in 21, exceptionally 23 rows | 2 |
| 1c. | Midbody dorsal scales arranged in 19 rows | 3 |
| 2a. | Large brownish dark-edged roundish, oval or saddle-shaped dorsal markings extending down both flanks, 118– 27 subcaudals | gracilis |
| 2b. | Two clearly demarcated slanting rougly “∏” shaped white markings on the back of head, 76–88 subcaudals | josephi sp. nov. |
| 3a. | Large brownish dark-edged oval or saddle-shaped dorsal markings extending down both flanks (nape element may be elongated and confined to middorsal segment, SE India) | bholanathi |
| 3b. | Dorsal colour pattern not as described (from western Nepal [unconfirmed] and northern India westward) | 4 |
| 4a. | With a reddish vertebral stripe | r. rhodorachis (partim) |
| 4b. | Vertebral stripe absent, postsubocular absent (two supralabials entering eye) or present (oftenso in mintonorum), dorsal colour pattern not as described except in certain juveniles of ventromaculatus | 5 |
| 5a. | Fewer than 220 ventrals and 332 ventrals plus subcaudals | 6 |
| 5b. | More than 219 ventrals and 331 ventrals plus subcaudals | 8 |
| 6a. | Nuchal streak present (very rarely absent), dorsal colour pattern along entire trunk | ventromaculatus |
| 6b. | Nuchal streak absent (very rarely present), last quarter of trunk devoid of pattern or unicoloured throughout | 7 |
| 7a. | 199 – 204 ventrals, 108 to ca. 112 subcaudals, sum 307 to ca. 314 (only ♂♂ known, probably close to and above 1,800 m a.s.l. east of Nushki, Urak Valley area, and Ziarat Mountains in NE Balochistan, Pakistan) | noeli |
| 7b. | More than 203 ventrals and 105 subcaudals, sum at least 318 (minima from Hindukush to N Khyber Pakhtunkhwa) | r. rhodorachis (partim) |
| 8a. | 110–127 subcaudals, postsubocular present or absent (0 – 2 supralabials entering eye), preocular often divided (northern Balochistan Region) | mintonorum |
| 8b. | 106–148 subcaudals (132–144 in r. rhodorachis from area of confirmed sympatry wit mintonorum), postsubocular usually absent (two supralabials entering eye), preocular entire (very rarely divided) | 9 |
| 9a. | 230–247 ventrals, 133–148 subcaudals, sum 363–395 (documented for Sindh and contiguous Balochistan Province, probably also SE Punjab and possibly W Rajasthan) | sindhensis |
| 9b. | 220–256 ventrals (fewer than 220 in area of potential sympatry with sindhensis), 106–145 subcaudals, sum usually lower than 364 (maximum in Makran 364, as much as 374 in NE Balochistan and 388 in Chagai) | 10 |
| 10a. | Usually fewer than 233 ventrals (as much as 244 in Chagai), 19 dorsal scale rows on anterior trunk or at midbody and 11–13 prior to vent | r. rhodorachis (partim) |
| 10b. | 237–256 ventrals, 19–21 dorsal scale rows on anterior trunk or at midbody and 13–15 prior to vent (eastern Hindu Kush, Karakoram, Ladakh, Spiti Valley in NE Himachal Pradesh, and probably Zanskar) | rhodorachis ladacensis |
Discussion
The maximum likelihood (ML) and Bayesian (BI) phylogeny presented in this study were similar but not same, there were some unresolved nodes in the BI phylogeny. Furthermore, the phylogenetic relationships are different from that presented by
Platyceps josephi sp. nov. is so far recorded only from Tamil Nadu state and all except two records (Salem and Auroville, Villupuram district) are south of the Cauvery River. The Nilgiri Mountains, the Moyar gorge and Cauvery River have been proposed to be a biogeographic barrier for fan-throated lizards (
In their study on Western Palearctic colubrid snakes,
Acknowledgements
We thank Sameresh Changdar, Vikash Behari and Parag Nag at GE Inspections Technologies, Chakan, Pune for their support and help in microCT scanning of the holotype of Joseph’s racer. We would like to thank Anil Khaire (Chairman Indian Herpetological Society, Pune), Vishal Verma, Sandeep Belkhede, Madhao Vaidya and Varad Giri (NCBS, Bangalore), V.J. Jins (SACON, Coimbatore), Chethan Kumar (Wildlife Institute of India, Dehradun) Raju Vyas (Gujarat) and Caleb Daniel (IISc, Bangalore), Suvendu Swain and Rakesh K. Mohalik (Odisha) for donating tissue samples for this study. We would like to thank Praveen Karanth for the lab facilities, Chinta Sidharthan and Kunal Arekar for support during lab work and Natalie Cooper for help with some of the analyses. We would also like to thank S.P. Vijayakumar for his help and inputs on microCT scanning. We are grateful to Naveen Joseph’s family and friends (Akila Joseph, Eswarapandi, Mahendraraja, Zakaria) for their support.
We are indebted to curators and other staff who approved loans made us welcome at their collections or provided photographs and information about specimens under their care used for this study. These are Rahul Khot, Saunak Pal, Mrugank Prabhu, Vithoba Hegde, Parvathy and Shyam Jadhav (
We are grateful to Rea Kavazi and Gillian Humphreys (Bodleian Library, Oxford, U.K.), David J. Gower (NHM London U.K.), John Rose, Andrea Hart and Rosie Jones (NHM Library, London, U.K.) who assisted us in the search for unpublished manuscripts and original drawings from T. E. Cantor, B. H. Hodgson and T. Hardwicke, and permitted us to reproduce them. For the support in procuring rare literature, we thank Hans-Ulrich Raake and Martina Rißberger (Zoological Main Library, Museum für Naturkunde Berlin, Germany), as well as Bernhard Schurian (Museum für Naturkunde Berlin, Germany) for scanning and postprocessing of the plates, reproduced from Russell`s book. Christoph Kucharzewski (
References
- Abercromby AF (1910) The snakes of Ceylon. Murray & Co., London, 89 pp.
- Abercromby AF (1913) Distribution of snakes in Ceylon. Spolia Zeylanica 8(21): 304–305.
- Abyerami S, Sivashanthini K (2008) Diversity of snakes from Jaffna Peninsula, Sri Lanka. Pakistan Journal of Biological Sciences 11(16): 1969–1978. https://doi.org/10.3923/pjbs.2008.1969.1978
- Adler K (2007) Cantor, Theodore (1809–1860). In: Adler K (Ed.) Herpetologists of the past. SSAR Contributions to the history of herpetology, Volume 2, St. Louis, New York, 65–66.
- Adler K (2016) Theodore Cantor (1809–1860): Pioneer scientific herpetologist in China [Introduction]. In: Adler K (Ed.) Theodore Cantor. Zoology of Chusan. SSAR Reprint, Ithaca, New York, v–xi.
- Aengals R (2009) Reptilia. In: Zool. Surv. India (Ed.): Fauna of Tamil Nadu, (Part-1). Zoological Survey of India, State Fauna Series 17, Kolkata, 187–198.
- Aengals R, Pradhan MS (2013) Reptiles. In: Zool. Surv. India (Ed.) Fauna of Karnataka. Zoological Survey of India, State Fauna Series 21, Kolkata, 435–465.
- Aengals R, Kumar VMF, Palot MJ, Ganesh SR (2018) A checklist of Reptiles of India. Zoological Survey of India, (Fauna of India Checklist), Kolkata, 35 pp.
- Agrawal HP (1976) Fauna of Kanha National Park – Reptilia. Newsletter of the Zoological Survey of India 2(6): 247–249.
- Agrawal HP (1979) A check-list of reptiles of Himachal Pradesh, India. The Indian Journal of Zootomy 20(2): 115–124.
- Ahmed M, Basak K, Sarkar P, Suraj M, Bebarta KC (2015) Snakes and other reptiles of Chhattisgarh. The field guide. State Biodiversity Board, Raipur, 212 pp.
- Ahmed S, Dasgupta G (1992) Reptilia. In: Gosh (Ed.) Fauna of West Bengal. Part 2. – Zoological Survey of India, State Fauna Series 3, Calcutta, 1–65.
- Amaral A do (1929) Estudos sobre Ophidios Neotropics. XVII. Valor systematico de varias formas de Ophidios Neotropics. Memorias do Instituto Butantan 4: 3–68.
- Amol K, Anup P, Gaurang P, Anant P, Dipankar L (2012) Notes on observed snakes in Satpura Tiger Reserve and record of Psammophis longifrons Boulenger, 1896 Madhya Pradesh, India. Universal Journal of Environmental Research and Technology 2(4): 362–365.
- Amr ZS, Disi AM (2011) Systematics, distribution and ecology of the snakes of Jordan. Vertebrate Zoology 61(2): 179–266.
- Anders CC, Kästle W, Shah KB, Schleich HH (1997) Die Amphibien und Reptilien Nepals. In: Schleich HH (Ed.) Das andere Nepal. Eine Ausstellung im Fuhlrott Museum Wuppertal. Fuhlrott-Museum Wuppertal, 41–54.
- Anderson J (1871) A list of the reptilian accession to the Indian Museum, Calcutta, from 1865 to 1870, with a description of some new species. Journal of the Asiatic Society of Bengal (Part II) 40(1): 12–39.
- Anderson J (1879) Anatomical and zoological researches: Comprising an account of the zoological results of the two expeditions to Western Yunnan in 1868 and 1875 […]. First Volume. Bernard Quaritch, London, [1878], xxv+984+(1) pp.
- Anderson JA (1964) A report on the Gecko Teratolepis fasciata (Blyth, 1853). Journal of the Bombay Natural History Society 61(1): 161–171.
- Anonymous (n.d.) Ecological profile Pratapgarh, Rajasthan. Foundation for Ecological Security, Anand, 30 pp.
- Anonymous (1796) [? Gmelin, F. J.] An account of Indian serpents collected on the coast of Coromandel, […]. In: Göttingische Anzeigen von gelehrten Sachen unter Aufsicht der königl. Gesellschaft der Wissenschaften. 167. Stück. Den 17. October 1796. Göttingische Anzeigen [etc.], Der zweyte Band auf das Jahr 1796, 1665–1670 [review of Russell (1796)].
- Anonymous (1822) Tentamen Systematics Amphibiorum. Auctore Blasio Merrem. Isis von Oken 1(4): col. 688–704. [review of Merrem (1820)].
- Anonymous (1870) Accessions to the Museum in July 1870 II. Reptilia. Indian Museum. Annual Report and List of Accessions 1870: 43–46.
- Anonymous (1877) Accessions to the Indian Museum from 1st April 1876 to 31st March 1877. Reptilia, Amphibia. Indian Museum. Annual Report and List of Accessions 1877: 40–45.
- Anonymous (1879) Accessions to the Indian Museum during the quarter ending 31st December 1878. Reptilia, Batrachia. Indian Museum. Annual Report and List of Accessions 1879: 35.
- Anonymous (2010) Defending Biodiversity and Promoting Sustainable Agriculture in Jalgaon District (India) Integrating New Methodologies and Advanced Technologies: Hotspot Geoinformatics, Remote Sensing, Geographical Information Systems, Global Positioning Systems and Smart Ground Sensors. Jala SRI Watershed Surveillance and Research Institute, Jalgaon (Maharashtra, India), Project Report, (3)+viii+150 pp.
- Anonymous (2017) Environmental Management Plan for Laterite Stone Quarries: Lingdal Cluster, Taluka: Devgad, Dist: Sindhudurg, Maharashtra. Sadekar Enviro Engineers Pvt. Ltd., 100+(12)+12 (Annex) pp.
- Arévalo E, Davis SK, Sites JW (1994) Mitochondrial DNA sequence divergence and phylogenetic relationships among eight chromosome races of Sceloporus grammicus complex (Phrynosomatidae) in central Mexico. Systematic Biology 43: 387–418. https://doi.org/10.1093/sysbio/43.3.387
- Auffenberg W (1963) The fossil snakes of Florida. Tulane Studies in Zoology 2: 89–155.
- Bachman S, Moat J, Hill AW, De La Torre J, Scott B (2011) Supporting Red List threat assessments with GeoCAT: geospatial conservation assessment tool. ZooKeys 150: 117. https://doi.org/10.3897/zookeys.150.2109
- Bauer AM (2015) Patrick Russell’s snakes and their role as type specimens. Hamadryad 37(1&2): 18–65.
- Bauer AM, Vogel G, Campbell PD (2015) A preliminary consideration of the dry snakes skin specimens of Patrick Russell. Hamadryad 37(1&2): 73–84.
- Bahuguna A (2010) Reptilia. In: Zool. Surv. India (Ed.): Fauna of Uttarakhand, (Part 1), Vertebrates. Zoological Survey of India, State Fauna Series 18, Kolkata, 445–503.
- Bansode SA, More VR (2018) An updated list of serpents from Palghar district of Maharashtra, India. Journal of Entomology and Zoology Studies 6(1): 375–379.
- Bansode SA, More VR, Mirza KA (2016) A Study on snakes from Mokhada and Jawhar (Dist. Palghar) Maharashtra, India. International Journal of Fauna and Biological Studies 3(5): 103–115.
- Bechstein JM (1802) Herrn de la Cepede’s Naturgeschichte der Amphibien oder der eyerlegenden vierfüßigen Thiere und der Schlangen. Eine Fortsetzung von Buffon’s Naturgeschichte. Aus dem Französischen übersetzt und mit Anmerkungen und Zusätzen versehen. Vierter Band. Industrie-Comptoir, Weimar, xx+298 pp., 48 pls.
- Beddome RH (1862) Notes upon the land and fresh-water snakes of the Madras presidency. The Madras Quarterly Journal of Medical Science 5: 1–31, pl. 2.
- Bethencourt Ferreirea J (1897) Reptis da India no Museu de Lisboa. Journal de Sciencias Mathematicas, Physicas e Naturaes da Academia Real das Sciencias de Lisboa Ser. 2, 4(16): 212–234.
- Bhandarkar WR, Paliwal GT, Bhandarkar SV, Kali AA (2012) Herpetofaunal diversity at Navegaon National Park, Distt. Gondia Maharashtra. International Journal for Environmental Rehabilitation and Conservation 3(1): 42–49.
- Bhupathy S, Sathishkumar N (2013) CEPF Western Ghats special series: status of reptiles in Meghamalai and its environs, Western Ghats, Tamil Nadu, India. Journal of Threatened Taxa 5(15): 4953–4961. https://doi.org/10.11609/JoTT.o3595.4953-61
- Bhuvaneswari R (2003) Pulicat Lake. In: Bioresources status in selected coastal locations. National Biorecources Development Board, Department of Biotechnology, Govt. of India, New Delhi and M.S. Swaminathan Research Foundation, Chennai, 142–176.
- Blanchflower P (2005) Restoration of the tropical dry evergreen forest of peninsular India. Biodiversity, 6(1), 17–24. https://doi.org/10.1080/14888386.2005.9712755
- Blanford WT (1879) Notes on a collection of reptiles and frogs from the neighbourhood of Ellore and Dumagudem. Journal of the Asiatic Society of Bengal (Part II) 48(12): 110–116.
- Blyth E (1854) Report of the Curator. Zoological Department. Journal of the Asiatic Society of Bengal [1853] 22(4): 408–417.
- Blyth E (1855a) Notices and descriptions of various reptiles, new or little known [part 2]. Journal of the Asiatic Society of Bengal [1854] 23(3): 287–302.
- Blyth E (1855b) Report of Curator, Zoological Department, for September, 1854. In: Proceedings of the Asiatic Society of Bengal for October, 1854. Journal of the Asiatic Society of Bengal [1854], 729–740.
- Blyth E (1860) Report of Curator, Zoological Department. Journal of the Asiatic Society of Bengal 29(1): 87–115.
- Boie F (1827) Bemerkungen über Merrem’s Versuch eines Systems der Amphibien. Marburg, 1820, 1te Lieferung, Ophidier. Isis von Oken 20(6): col. 508–566.
- Boie H (1826) Notice sur l’Erpétologie de l’Ile de Java. Bulletin Sci. Indust. Férussac (deuxième section Bull. Sci. Nat. Géol.) 9(2): 233–240 (edited by H. Schlegel).
- Boulenger G (1890) The Fauna of British India, including Ceylon and Burma. Reptilia and Batrachia. Taylor & Francis, London, xviii+541 pp.
- Boulenger G (1893) Catalogue of the snakes in the British Museum (Natural History). Volume 1. British Museum (Nat. Hist.), London, xiii+448 pp., pl. i–xxviii.
- Bourret R (1936) Les Serpents de l’Indochine. Tome II. Catalogue Systématique Descriptif. Henri Basuyau & Cie, Toulouse, 505 pp.
- Buhle CA, Thon T, Reichenbach AB (n.d., ca. 1835) Die Naturgeschichte in getreuen Abblildungen und mit ausführlicher Beschreibung derselben. Amphibien. C. Brüggemann and O. Wigand, Halberstadt and Pesth, (1)+100+(4) pp., 74 pls.
- Burmeister H (1850) Verzeichnis der im zoologischen Museum der Universität Halle-Wittenberg aufgestellten Säugethiere, Vögel und Amphibien. Zoologisches Museum Halle, 84 pp.
- Campbell PD (2015) The acquisition of Dr. Patrick Russell’s snake skin collections. Hamandryad 37(1&2): 66–72.
- Cantor T (1839) Spicilegium Serpentium Indicorum. Proceedings of the Zoological Society of London, vii: 31–34+49–55.
- Cantor T (1841) Spicilegium Serpentium Indicorum. The Calcutta Journal of Natural History 1: 76–79+80–86 [reprint of Cantor 1839].
- Cantor T (1847a) Catalogue of reptiles inhabiting the Malayan Peninsula and islands [parts 1–3]. Journal of the Asiatic Society of Bengal, new series 16: 607–656+897–952+1026–1078, 2 pls.
- Cantor T (1847b) Catalogue of reptiles inhabiting the Malayan Peninsula and islands [parts 1–3]. Calcutta (published by the author), 157 pp., 2 pls. [reissued 1886 in Trübner’s Oriental Series. Miscellaneous papers relating to Indo-China, Vol. II; reprinted 1966 by A. Asher & Co., Amsterdam, and by Cosmo Publictations, New Delhi in 1981].
- Cantor T (1848) Latitudinal distribution of reptiles inhabiting the Malayan Peninsula and islands, and other localities. The Edinburgh New Philosophical Journal 44: 271–286 [partly reprinted from Cantor (1847a, pt. 3)].
- Cardew AG (1897) A rough key to the identification of Indian ophidia. Journal of the Bombay Natural History Society 10(4): 585–596.
- Cazaly HW (1914) The common snakes of India and Burma and how to recognize them. Allahabad, The Pioneer Press, II+(1)+60+(13)+IV pp. [reprinted 1984 by International Book Distributors, Dehra Dun].
- Captain A, Thakur S (1999) Aberrant banded racers Argyrogena fasciolatus. Journal of the Bombay Natural History Society 96(1): 147–151.
- Chaitanya R, Khandekar A, Caleb DG, Mukherjee N, Ghosh A, Giri V (2018) Herpetofauna of the Meghamalai Wildlife Sanctuary, Southern Western Ghats, India: An updated checklist with annotations on taxonomy and nomenclature. Journal of the Bombay Natural History Society 115: 21–37.
- Chakrabarti P (2006) “Neither of meate nor drinke, but what the Doctor allowed”: Medicine amidst war and commerce in Eighteenth-Century Madras. Bulletin of the History of Medicine 80(1): 1–38.
- Chettri B, Acharya BK, Bhupathy S (2011) An overview of the herpetofauna of Sikkim with emphasis on the elevational distribution pattern and threats and conservation issues. In: Arrawatia ML, Tambe S (Eds.) Biodiversity of Sikkim: Exploring and conserving a global hotspot. Information and Public Relations Department, Government of Sikkim. Gangtok, 233–254.
- Chettri B, Bhupathy S (2007) Reptile fauna of Sikkim with emphasis to Teesta valley. Journal of Hill Research 20(1): 1–11.
- Chandra K, Gajbe PU (2005) An inventory of herpetofauna of Madhya Pradesh and Chhattisgarh. Zoo’s Print Journal 20(3): 1812–1819.
- Chandra K, Sharma RC, Nagpure A, Nema DK (2008) Reptiles of Kanha Tiger Reserve, Madhya Pradesh. Records of the Zoological Survey of India 108(4): 49–83.
- Charjan AP, Joshi PS (2015) Studies on diversity of snakes in Akola district of Maharashtra, India. Review of Research 4(6): 1–3.
- Chauhan RR, Shingadia HU (2012) Preliminary survey of herpetofauna of Borivali mangroves–a coastal belt in the suburbs of Mumbai. Life sciences Leaflets (LSIC2011): 59–65.
- Chikane S, Bhosale HS (2012) Die Reptilien von Kaas, nördliche Western Ghats, Maharashtra, Indien, mit Bemerkungen zu Vorzugsbiotopen, Häufigkeiten und Gefährdung. Sauria 34(3): 3–15.
- Cocker M, Inskipp C (1988) A Himalayan Ornithologist. The life and work of Brian Houghton Hodgson. Oxford University Press, Oxford, New York, Tokyo, xii+87 pp., 49 pls.
- Constable JD (1949) Reptiles from the Indian Peninsula in the Museum of Comparative Zoölogy. Bulletin of the Museum of Comparative Zoölogy 103(2): 59–160.
- Cope ED (1861) Catalogue of the Colubridae in the Museum of the Academy of Natural Sciences of Philadelphia. Part 3. Proceedings of the Academy of Natural Sciences of Philadelphia 12 [1860]: 553–566.
- Cope ED (1863) Notes upon some reptiles of the Old World. Proceedings of the Academy of Natural Sciences of Philadelphia 14 [1862]: 337–344.
- Crawford DG (1914) A history of the Indian Medical Service 1600–1913. Volume 1. W. Thacker & Co., Thacker Spink & Co., London, Calcutta & Simla, xix+(3)+529 pp., 2 pls.
- Crawford DG (1916) Two Madras Medical lists. The Indian Medical Gazette 51(1): 1–7.
- Cuvier G, Duméril AMC (1818) Coleuvre, Coluber. Dictionnaire des sciences naturelles, [etc.], XI. Cos–Cris., 170–216.
- Daniel JC (1989) The book of Indian reptiles. Bombay Natural History Society, Oxford University Press, 141 pp.
- Daniel JC (2002) The book of Indian reptiles and amphibians. Bombay Natural History Society, Oxford Univiversity Press, Mumbai, Delhi, Calcutta, Chennai, vii +238 pp.
- Daniels RJR (2000) Reptiles and amphibians of Karnataka. Cobra 42: 1–11.
- Daniels RJR (2001a) Amphibians and reptiles of the Tamil Nadu Eastern and Western Ghats. Cobra 43: 1–8.
- Daniels RJR (2001b) Snakes of Tamil Nadu: a status report. Cobra 44: 11–17.
- Das I (1994) The reptiles of South Asia: Checklist and distributional summary. Hamadryad 19: 15–40.
- Das I (1996) Biogeography of the reptiles of South Asia. Krieger Publ. Co., Malabar, vii+87 pp., 16 pls.
- Das I (1997) Checklist of the reptiles of India with English common names. Hamadryad 22(1): 32–45.
- Das I (1998) The serpent’s tongue. A contribution to the ethnoherpetology of India and adjacent countries. Edition Chimaira, Frankfurt am Main, 121 pp.
- Das I (2002) A photographic guide to snakes and other reptiles of India. New Holland Publ., London, 144 pp. [reprinted 2012].
- Das I (2003) Growth of knowledge on the reptiles of India, with an introduction to systematics, taxonomy and nomenclature. Journal of the Bombay Natural History Society 100(2&3): 446–501.
- Das I (2012) Reptiles. In: Dutta, S. K. (Ed.) Compendium on the occasion of DST 5th School in Herpetology, 24th February–10th March, 2012. North Orissa University, Baripada, Odisha, 63–80, pl.
- Das I, Andrews H (1997) Checklist of Indian Reptiles. In: Conservation Assessment and Management Plan (C.A.M.P.) Workshop for Reptiles of India, 19-23 May 1997, Coimbatore, 1–4.
- Das I, Ota H (1998) A checklist of chromosome numbers of south Asian reptiles. Hamadryad 23(2): 179–193.
- Das I, De Silva A (2005) A photographic guide to snakes and other reptiles of Sri Lanka. New Holland Publ. Ltd., London, 144 pp.
- Das I, Das A (2017) A naturalist’s guide to the reptiles of India, Bangladesh, Bhutan, Nepal, Pakistan and Sri Lanka. John Beaufoy Publishing, Oxford, 176 pp.
- Das I, Dattagupta B, Gayen NC (1998) History and catalogue of reptile types in the collection of the Zoological Survey of India. Journal of South Asian Natural History 3(2): 121–172.
- Das S, Pramanick K (2019) Comparative anatomy and homology of jaw adductor muscles of some South Asian colubroid snakes (Serpentes, Colubriodea). Vertebrate Zoology 69(1): 93–102. https://doi.org/10.26049/VZ69-1-2019-04
- Das S, Campbell PD, Roy S, Mukherjee S, Pramanick K, Biswas A, Raha S (2019) Cranial osteology and molecular phylogeny of Argyrogena fasciolata (Shaw, 1802) (Colubridae: Serpentes). Vertebrate Zoology 69(3): 311–325. https://doi.org/10.26049/VZ69-3-2019-04
- Dasgupta G, Raha S (2004) Reptilia. In: Zoological Survey of India (Ed.) Fauna of Bihar (including Jharkhand), Part 1. Zoological Survey of India, State Fauna Series 11, Kolkata, 143–179.
- Dasgupta G, Murthy BHCK, Raha S (2012) Reptilia. In: Zoological Survey of India (Ed.) Fauna of Maharashtra, Part 1, Vertebrates. Zoological Survey of India, State Fauna Series 20, Kolkata, 189–238.
- Daudin FM (1803) Histoire Naturelle, Génerale et Particulière des Reptiles; Ouvrage faisant suite aux Oeuvres de Leclerc de Buffon, et Partie du Cours Complet d’Histoire Naturelle Rédigé par C.S. Sonnini, membre de plusieurs sociétés savantes. Tome sixième. F. Dufart, Paris, 447 pp., pls. lxxi–lxxx.
- Deepak V, Karanth KP (2018) Aridificaton driven diversification of fan-throated lizards from the Indian subcontinent. Molecular Phylogenetics and Evolution 120: 53–62. https://doi.org/10.1016/j.ympev.2017.11.016
- Deranyiagala PEP (1955) A coloured atlas of some vertebrates from Ceylon. Volume three. Serpentoid reptilia. Government Press, Colombo, vii+(1)+49 pls. +(5)+121 pp.
- De Silva A (1990) Colour guide to the snakes of Sri Lanka. R & A Publishing Limited, Portishead, GB, 130 pp.
- De Silva A (1994) An introduction to the herpetofauna of Sri Lanka. Lyriocephalus 1(1/2): 3–19.
- De Silva A (1998a) Country report for Sri Lanka. Herpetofauna of Sri Lanka: Present status, distribution and conservation. In: de Silva A (Ed.) Proceedings of the International Conference on the Biology and Conservation of Amphibians and Reptiles of South Asia, Sri Lanka, August 1-5, 1996. Amphibia and Reptile Research Organization of Sri Lanka (ARROS), Peradeniya, 51–73.
- De Silva A (1998b) Snakes of Sri Lanka: A checklist and an annotated bibliography. Department of Wildlife Conservation/GEF/UNDP/FAO, Battaramulla, 6+112 pp.
- De Silva A (2001) The herpetofauna of Sri Lanka. An historical overview, current status with checklist. Amphibia and Reptile Research Organisation (ARROS), Peradeniya, (2)+91 pp.
- De Silva A, Jinasena J (2009) Snakes of Sri Lanka. A coloured atlas (With accounts on snakes in archaeology, history, folklore, venomous snakes and snakebite). Vijitha Yapa Publications, Colombo, xix+(1)+129+(1) pp.
- De Silva A, Ukuwela K (2017) A naturalist’s guide to the reptiles of Sri Lanka. John Beaufoy Publishing, Oxford, 176 pp.
- De Silva PHDH (1980) Snake fauna of Sri Lanka with special reference to skull, dentition and venom in snakes. National Museum of Sri Lanka, Colombo xi+472 pp.
- Deshmukh RV, Deshmukh SA, Badekar SA (2015) Rescued records of snakes from Nagpur district, Maharashtra with data on unrecorded species. Reptile Rap 17: 34–43.
- Dileepkumar R, Sharma VR, Oommen OV, Laladhas KP, Krishnan AS, Kumar AV (2016) Handbook on snakes of Kerala. Kerala State Biodiversity Board, Thiruvananthapuram, (12)+1–32+1–8+203+(1) pp.
- Dodwell [E], Miles [JS] (1838) (Eds) Alphabetical list of the officers of the Indian Army; with the dates of their respective promotion, retirement, resignation, or death, whether in India or in Europe; from the year 1760, to the year 1834 inclusive, corrected to September 30, 1837. London, Longman, Orme, Brown, and Co., 609 pp.
- Dowling HG (1951a) A proposed standard system of counting ventrals in snakes. British Journal of Herpetology 1(5): 97–99.
- Dowling HG (1951b) A proposed method of expressing scale reductions in snakes. Copeia 1951(2): 131–134.
- Dowling HG, Savage JM (1960) A guide to the snakes hemipenis: a survey of basic structure and systematic characteristics. Zoologica 45: 17–31.
- Druitt R (1860) The principles and practice of modern surgery. Blanchard and Lea, Philadelphia, xxiii+695+32 pp.
- Duméril AMC, Bibron G, Duméril A (1854) Erpétologie générale ou histoire naturelle complète des reptiles. Tome septième. Première partie. Roret, Paris, xvi+780 pp.
- Dundee HA (1994) Current names for Linnaeus’s herpetological species in Systema Naturae X. In: Kitchell K jr, Dundee HA. A trilogy on the herpetology of Linnaeus’s Systema Naturae X. – Smithsonian Herpetological Information Service, Washington 100, 41–61.
- Dunkan A sen, Duncan A jun (1798) Russell, Patrick, M.D.F.R.S. An Account of Indian Serpents collected on the coast of Coromandel; [etc.]. Annals of Medicine, for the Year 1797 [Sect. I. Analysis of Books], Edinburgh 2: 1–18.
- Dunn ER (1928) A tentative key and arrangement of the American genera of Colubridae. Bulletin of the Antivenin Institute of America 2(1): 18–24.
- Dutta SK, Acharjyo LN (1990) Checklist of the herpetofauna of Orissa, India. The Snake 22: 36–43.
- Dutta SK, Nair MV, Mohapatra PP, Mahapatra AK (2009) Amphibians and reptiles of Similipal biosphere reserve. Government of Odisha, Regional Plant Resource Centre publication, Bhubaneswar, 174 pp.
- Fellows S (2014) Species Diversity of Snakes in Pachmarhi Biosphere Reserve. Entomology, Ornithology & Herpetology: Current Research 4(1): 1–4. https://doi.org/10.4172/2161-0983.1000136
- Ferguson W (1877) Reptile fauna of Ceylon. Letter on a collection sent to the Colombo Museum. William Henry Herbert, Govt. Printer, Colombo, 42 pp. [reprinted 2001 by the Society for the Study of Amphibians and Reptiles, Ithaca, New York].
- Figueroa A, McKelvy AD, Grismer LL, Bell CD, Lailvaux SP (2016) A species-level phylogeny of extant snakes with description of a new colubrid subfamily and genus. PloS One 11(9): 1–31. https://doi.org/10.1371/journal.pone.0161070
- Filippi F de (1840) Catalogo ragionato e descrittivo della Raccolta de’ Serpenti del Museo dell’ I. R. Università di Pavia. Biblioteca Italiana 99: 163–187+306–343 [reissued 1840 as authors reprint, 65 pp.].
- Fitzinger LJ [“I”] (1826) Neue Classification der Reptilien nach ihren natürlichen Verwandtschaften. Nebst einer Verwandtschafts-Tafel und einem Verzeichnisse der Reptiliensammlung des k. k. zoologischen Museum’s zu Wien. J. G. Heubner, Wien, [6]+66 pp.
- Fitzinger L [J] (1843) Systema Reptilius. Fasciculus Primus Amblyglossae. Vindobonae [Vienna], Braumüller et Seidel, 106+vi+3 pp. [reprinted 1973 by the Society for the Study of Amphibians and Reptiles, Oxford, Ohio].
- Felsenstein J (1981) Evolutionary trees from DNA sequences: a maximum likelihood approach. Journal of Molecular Evolution 171: 368–376. https://doi.org/10.1007/BF01734359
- Fraser A (1936) The Snakes of Deolali. With notes on their comparative osteology and peculiarities of dentition. Part 1–3. Journal of the Bombay Natural History Society 39(1): 58–82, 36(2): 264–291, 36(3): 464–501.
- Fricke R, Eschmeyer WN, Van der Laan R (Eds) (2020) Eschmeyer’s Catalog of Fishes: Genera, Species, References. (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp). Electronic version accessed 06.04.2020.
- Froriep R, Schomburgk O (1848) Amphibien der malaiischen Halbinsel, wie überhaup6 der Inseln des Süd-Oceans. Nach Cantor. Fortschritte der Geographie und Naturgeschichte Vol. 5, 69(1): col. 279–284.
- Gajbe P, Gupta R (2005) Herpetofauna of Shahdol District, Madhya Pradesh. Reptile Rap 7: 9–10.
- Ganesh SR, Asokan JR (2010) Catalogue of Indian herpetological specimens in the collection of the Government Museum Chennai, India. Hamadryad 35(1): 46–63.
- Ganesh SR, Chandramouli SR (2011) Report of some noteworthy specimens and species of Herpetofauna from South-East India. Taprobanica 3(1): 5–10. http://doi.org/10.4038/tapro.v3i1.3227
- Ganesh SR, Vogel G (2018) Taxonomic reassessment of the Common Indian Wolf Snake Lycodon aulicus (Linnaeus, 1758) complex (Squamata: Serpentes: Colubridae). Bonn zoological Bulletin 67(1): 26–36.
- Ganesh SR, Kalaimani A, Karthik P, Baskaran N, Nagarajan R, Chandramouli SR (2018) Herpetofauna of southern Eastern Ghats, India – II from Western Ghats to Coromandel Coast. – Asian Journal of Conservation Biology 7(1): 28–45.
- Ganesh SR, Deuti K, Punith KG, Achyuthan NS, Mallik AK, Adhikari O, Vogel G (2020) A new species of Lycodon (Serpentes: Colubridae) from the Deccan Plateau on India, with notes on the range of Lycodon travancoricus (Beddome, 1870) and a revised key to peninsular Indian forms. Amphibian & Reptilie Conservation [Taxonomy Section] 14(3): 74–83.
- Gayen D, Dey S, Roy US (2017) Diversity of snakes in and around Durgapur city, West Bengal, India. Reptile Rap 176, in: Zoo’s Print 32(9): 17–22.
- Gayen N (1999) A synopsis of the reptiles of Gujarat, western India. Hamadryad 24(1): 1–22.
- Gemel R, Gassner G, Schweiger S (2019) Katalog der Typen der Herpetologischen Sammlung des Naturhistorischen Museums Wien–2018. Annalen des Naturhistorischen Museums Wien B, 121: 33–248.
- Georgi JG (1801) Geographisch-physikalische und naturhistorische Beschreibung des Russischen Reichs, zur Uebersicht bisheriger Kenntnisse von demselben. Des dritten Theils siebenter Band. Friedrich Nicolovius, Königsberg 7(3), (1)+1681–2222 pp.
- Gharpurey KG (1931) An unusually large Shaw’s Rat Snake (Zamenis fasciolatus). Journal of the Bombay Natural History Society 34(4): 1084.
- Gharpurey KG (1932a) Number of the ventral scales in the Fasciolated Dhaman (Z. fasciolatus). Journal of the Bombay Natural History Society 35(2): 465.
- Gharpurey KG (1932b) Note on the Fasciolated Rat Snake (Zamenis fasciolatus). (A correction) Journal of the Bombay Natural History Society 35(4): 906.
- Gharpurey KG (1932c) Snakes in Ahmednagar. Journal of the Bombay Natural History Society 36(1): 272–273.
- Gharpurey KG (1935a) Snakes in Belgaum. Journal of the Bombay Natural History Society 37(4): 942–944.
- Gharpurey KG (1935b) A further list of snakes from Ahmednagar. Journal of the Bombay Natural History Society 38(1): 198–200.
- Gmelin JF (1789) Caroli a Linné. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio Decima Tertia, Aucta, Reformata. Tom. I Pars III. Lipsiae, Georg Emanuel Beer, (1)+1033–1516 pp.
- Gmelin JF (1798) Auszug aus Patr. Russell’s Nachricht von indischen Schlangen. Göttingisches Journal der Naturwissenschaften 1(3): 1–54.
- Gmelin JF (1809) A genuine and universal system of natural history; comprising the three kingdoms of animals, vegtetables, and minerals, arranged under their respective classes, orders, genera, and species by the late Sir Charles Linnaeus […] improved, corrected and enlarged. Volume the Twelfth [Reptiles]. W. Lewis, London, 735 pp., 21 pls.
- Gray JE (1831) A synopsis of the species of the class Reptilia. In: Griffith, E. and Pidgeon, E. (Eds) The animal kingdom arranged in conformity with its organization, by the Baron Cuvier, member of the Institute of France, &c. &c. &c. With additional descriptions of all the species hitherto named, and of many not before noticed. Volume the ninth. The class Reptilia arranged by the Baron Cuvier, with specific descriptions. Whittaker, Treacher, London, 1–110 (Appendix).
- Gray JE (1834) Illustrations of Indian Zoology; chiefly selected from the collection of Major-General Hardwicke. Vol. II. Adolphs Richter & Co. and Parbury, Allen & Co., London, 31 plates [no. 57–87 of reptiles].
- Gray JE (1863) Catalogue of the specimens and drawings of mammals, birds, reptiles and fishes of Nepal and Tibet, presented by B. H. Hodgson, ESQ., to the British Museum. British Museum (Nat. Hist.), London, second edition, xii+90 pp.
- Green HW (1988) Antipredator Mechanism in Reptiles. In: Gans C, Huey RB (Eds) Biology of the reptilia. Volume 16, Ecology B, Defense and life history. Alan R. Liss, Inc., New York, 1–152.
- Groth JG, Barrowclough GF (1999) Basal divergences in birds and the phylogenetic utility of the nuclear RAG-1 Gene. Molecular Phylogenetics & Evolution. 12: 115–123. https://doi.org/10.1006/mpev.1998.0603
- Gruber U (2002) Family Colubridae (Colubrid snakes). In: Schleich HH, Kästle W (Eds) Amphibians and reptiles of Nepal. A.R.G. Gantner K.G., Ruggell, 803–950.
- Günther A[CLG] (1858) Catalogue of colubrine snakes in the collection of the British Museum. Printed by order of the Trustees, London, xvi+281 pp. [reprinted 1971 by Alden Press, Oxford].
- Günther A[CLG] (1860) Contributions to a knowledge of the reptiles of the Himalaya mountains. Proceedings of the Zoological Society of London [1860]: 148–175, pl. xxv–xxviii.
- Günther A[CLG] (1861) List of the cold-blooded vertebrata collected by B. H. Hodgson, Esq., in Nepal. Proceedings of the Zoological Society of London [1861]: 213–227.
- Günther A[CLG] (1862) On new species of snakes in the collection of the British Museum. The Annals and Magazine of Natural History, ser. 3, 9(50): 124–132.
- Günther A[CLG] (1864) The reptiles of British India. R. Hardwicke [Ray Society], London, xxvii+452 pp. [reprinted 198? by Oxford & IBH Publ., New Delhi].
- Günther A[CLG] (1866) Fifth account of new species of snakes in the collection of the British Museum. The Annals and Magazine of Natural History, ser. 3, 18(103): 24–29.
- Günther A[CLG] (1875) Second report on collections of Indian reptiles obtained by the British Museum. Proceedings of the Zoological Society of London [1875]: 224–234.
- Haly A (1886) First report on a collection of snakes in the Colombo Museum. Presented by the Committee of the Colombo Museum, 18 pp.
- Haly A (1891) Report on the collection of reptilia & batrachia in the Colombo Museum. The Government Printing Works, Colombo, 39 pp.
- Hasan MK, Khan MMH, Feeroz MM (2014) Amphibians and reptiles of Bangladesh. A field guide. Arannayk Foundation, Dhaka, 191 pp.
- Hemming F, Noakes D (Ed.) (1958) Official list of works approved as available for zoological nomenclature. First installment: titles 1–38. London, The International Trust for Zoological Nomenclature, x+(1)+1–12.
- Hoeven J van der (1856) Handbuch der Zoologie. Zweiter Band (Wirbelthiere). Leopold Voss, Leipzig, x+817+(3) pp., 6 pls.
- Huelsenbeck JP, Ronquist F, Nielsen R, Bollback JP (2001) Bayesian inference of phylogeny and its impact on evolutionary biology. Science 294: 2310–2314. http://doi.org/10.1126/science.1065889
- Hutton AF (1949) Notes on the snakes and mammals of the High Wavy Montains, Madura district, S. India. Part I–Snakes. Journal of the Bombay Natural History Society 48(3): 453–460.
- Hutton AF, David P (2008) Note on a collection of snakes from South India, with emphasis on the snake fauna of the Meghamalai Hills, (High Wavy Mountains). Journal of the Bombay Natural History Society 105(3): 299–316.
- IUCN [International Commission on Zoological Nomenclature] (1958) Opinion 525. Determination of the relative precedence to be accorded to the names “Coluber pictus” Gmelin (J.F.), [1789], and “Coluber boiga” Lacépède, 1789 (Class Reptilia). Opinions and Declarations rendered by the International Commission on Zoological Nomenclature 19(12): 277–290.
- IUCN [International Commission on Zoological Nomenclature] (1999) International code of zoological nomenclature (4th ed.). The International Trust for Zoological Nomenclature, London, xxix+306 pp.
- IUCN [International Commission on Zoological Nomenclature] (2012) IUCN Red List Categories and Criteria, Version 3.1. (second edition). IUCN, Switzerland and United Kingdom.
- Ingle M (2001a) Notes on reptiles from Ujjain. Cobra 43: 14–15.
- Ingle M (2001b) Snakes of Madhya Pradesh, Ujjain and Malwa region. Cobra 45: 22–24.
- Ingle M (2002) Ecology and status of the ophiofauna of eight districts of Malwa region of Madhya Pradesh. Cobra 50: 1–17.
- Ingle M (2003) A preliminary survey of the herpetofauna of Naglok area, Jashpur, Chhatisgarh state. Cobra 54: 1–5.
- Ingle M (2008) Herpetofaunal diversity of Ujjain district, Madhya Pradesh. Cobra [new series] 2(2): 10–16.
- Ingle M (2011) Herpetofauna of Naglok region, Jashpur District, Chhattisgarh. Records of the Zoological Survey of India 111(4): 79–96.
- Ingle M, Yadav U, Bhilala AK, Pawar M (2019) Urban Herpetofauna: A case study in Ujjain City of Central India. International Journal of Scientific Research in Biological Sciences 6(5): 76–84.
- Ingle P, Bali S, Khandagale J (2014) Preliminary Survey of Snake Diversity from Malegaon Tehsil of Washim District. World Journal of Zoology 9(2): 134–137.
- Iskandar D, Colijn E (2002) A checklist of Southeast Asian and New Guinean reptiles. Part I: Serpentes. Department of Biology, Institute of Technology, Bandung, (1)+(1)+195 pp.
- Islam MA, Ameen M-U, Nishat A (Eds) (2000) Red book of threatened amphibians and reptiles of Bangladesh. Dhaka IUCN - The World Conservation Union, Dhaka, xii+95 pp.
- Jadhav PL, Chavan SP, Trimukhe HS (2018) Snake species diversity and their distribution in and around Nanded city, Maharashtra, India. Journal of Entomology and Zoology Studies 6(4): 1855–1860.
- Jan G (1863) Elenco sistematico degli ofidi descritti e disegnati per l’Iconografia generale. A. Lombardi, Milano, vii+143 pp.
- Jerdon TC (1853) Catalogue of reptiles inhabiting the peninsula of India. Journal of the Asiatic Society of Bengal 22(5): 462–479, (6): 522–534.
- Jha A, Thapa K (2002) Reptiles & amphibians of Sikkim. Mrs. S. Jha Publ., Raipur, 6+100 pp.
- Joshi PS (2011) A preliminary survey on the snakes of Buldhana district, Maharashtra. Golden Research Thoughts, 1(2): 1–2.
- Joshi PS, Tantarpale VT, Kulkarni KM (2014) A review of ophidian studies in Vidarbha region (M.S.) India. Science Research Reporter 4(2): 167–170.
- Joshi PS, Tantarpale VT, Kulkarni KM (2015) Seasonal diversity and population dynamics of ophidian fauna in Buldhana district Maharashtra India. Indian Journal of Science Research 6(1): 23–28.
- Joshi PS, Charjan AP, Tantarpale VT (2017) A Herpetofaunal Inventory of Vidarbha Region, Maharashtra, India. Bioscience Discovery 8(3): 582–587.
- Kabir SMH, Ahmad M, Ahmed ATA, Rahmann AKA, Begum ZNT, Hassan MA, Khondker M (Eds) (2009) Encyclopedia of Flora and Fauna of Bangladesh, Vol. 25. Amphibians and Reptiles. Asiatic Society of Bangladesh, Dhaka, xviii+204 pp.
- Kästle W, Rai K, Schleich HH (2013) Field guide to amphibians and reptiles of Nepal. ARCO-Nepal, Jhapa, 609 pp. [bilingual English/Nepali].
- Kalki Y, Gowda S, Agnivamshi M, Singh K, Patel H, Mirza ZA (2020) On the taxonomy and systematics of the recently described Lycodon deccanensis Ganesh, Deuti, Punith, Achyuthan, Mallik, Adhikari, Vogel, 2020 (Serpentes, Colubridae) from India. Evolutionary Systematics 4: 109–118. https://doi.org/10.3897/evolsyst.4.60570
- Kanaujia A, Kumar A, Kumar A (2017) Herpetofauna of Uttar Pradesh, India. Biological Forum-An International Journal 9(1): 118–130.
- Keogh JS (1999) Evolutionary implications of hemipenial morphology in the terrestrial Australian elapid snakes. Zoological Journal of the Linnean Society 125: 239–278. https://doi.org/10.1006/zjls.1997.0163
- Khaire N (1996) Indian Snakes. Indian Herpetological Society, Pune, 106 pp.
- Khaire N (2008) A guide to the snakes of Maharashtra Goa & Karnataka. Indian Herpetological Society `USANT’, Pune, 129 pp. [second edition].
- Khaire N (2010) Snakes. M. L. Paranjape & J. Prakashan Publ., Pune, 103+(1) pp. (third reprint 2013), [in Maharati].
- Khaire N (2014) Indian snakes. A field guide. Jyotsna Prakashan, Pune, 160 pp.
- Khan BN, Ahmad R, Ali Z, Yasmeen R, Azhar M, Abid F, Mehmood S, Raza H (2018) Study of vertebrate diversity at Lal Suhanra National Park, Pakistan. The Journal of Animal & Plant Sciences 28(6): 1725–1734.
- Khan MAR (1988) An updated list of snakes of Bangladesh. Hamadryad 13(1): 5–7.
- Khan MAR (2004) Checklist of the herpetofauna of Bangladesh. Cobra 57: 1–31.
- Khan MS (1977) Checklist and key to the snakes of Jhang District, Punjab, Pakistan. Biologia 23(2): 145–157.
- Khan MS (1980) Affinities and zoogeography of Herpetiles of Pakistan. Biologica 26(1–2): 113–171.
- Khan MS (1982) Some notes on snakes in Pakistan. Hamadryad 7(1): 4–5.
- Khan MS (1993) Sarzameen-a-Pakistan kay Saamp [Snakes of Pakistan]. Urdu Science Board, 208 pp. [in Urdu].
- Khan MS (1999) Herpetology of habitat types of Pakistan. Pakistan Journal of Zoology 31(3): 275–289.
- Khan MS (2002) Die Schlangen Pakistans. Edition Chimaira, Frankfurt am Main, 165 pp.
- Khan MS (2004) Annotated checklist of amphibians and reptiles of Pakistan. Asiatic Herpetological Research 10: 191–201.
- Khan MS (2006) Amphibians and reptiles of Pakistan. Krieger Publishing Company, Malabar, FL, xvi+311 pp.
- Khan MS (2016) An up to date checklist of reptiles of Pakistan. Pakistan Journal of Wildlife, 7(2): 21–27.
- Khan MZ, Ghalib SA, Siddiqui S, Siddiqui TF, Farooq RY, Yasmeen G, Abbas D, Zehra A (2012) Current status and distribution of reptiles of Sindh. Journal of Basic & Applied Sciences 8: 160–168.
- Khedkar T, Sharma R, Tiknaik A, Khedkar G, Naikwade BS, Ron TB, Haymer D (2014) DNA barcoding using skin exuviates can improve identification and biodiversity studies of snakes. Mitochondrial DNA A DNA Mapp Seq Anal 27(1): 499–507. https://doi.org/10.3109/19401736.2014.905830
- Koul O, Murphy JC (1978) The ecology of Indian amphibians and reptiles-A review. Bulletin of the Chicago Herpetological Society 14(1&2): 4–66.
- Koushik (2015) Saap. Naturism, Kolkata, 198 pp. [second edition].
- Kramer E. (1977) Zur Schlangenfauna Nepals. Revue suisse de zoologie 84(3): 721–761.
- Kuch U, Mebs D (2002) Envenomation by colubrid snakes in Africa, Europe and the Middle East. Journal of Toxicology-Toxin Reviews 21(1&2): 159–179. https://doi.org/10.1081/TXR-120004745
- Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870–1874. https://doi.org/10.1093/molbev/msy096
- Kumar PVR, Kameswaran S, Rajendran MV, Rajendran S, Kutty MN (1976) Response of certain fishes and snakes to sound. Journal of the Bombay Natural History Society 73(1): 88–93.
- Kumbhar A, Pradhan A, Patwardhan G, Pande A, Lahkar D (2012) Note on observed snakes in Satpura Tiger Reserve and record of Psammophis longifrons Boulenger, 1896 in Madhya Pradesh, India. Universal Journal of Environmental Research and Technology 2(4): 362–365.
- Kutty MN, Sukumaran N, Narayanan M (1981) Influence of ambient oxygen on respiratory quotient and survival of some South Indian snakes. Proceedings of the Indian national Science Academy B 47(2): 166–169.
- Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B (2016) PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Molecular Biology and Evolution 34(3): 772–773. https://doi.org/10.1093/molbev/msw260
- Laurenti JN (1768) Specimen medicum exhibens synopsin Reptilium emendatum cum experimentis circa venena et antidota Reptilium Austriacorum. Joan. Thomae nob. de Trattern, Viennae, (8)+214+(3) pp., 5 pls. [reprinted 1966 by A. Asher & Co., Amsterdam; facsimile reprint with an English translation by Sergius L. Kuzmin 2005, Laurenti Verlag, Bielefeld].
- Lawson R, Slowinski JB, Crother BI, Burbrink FT (2005) Phylogeny of the Colubroidea (Serpentes):new evidence from mitochondrial and nuclear genes. Molecular Phylogenetics & Evolution. 37: 581–601. https://doi.org/10.1016/j.ympev.2005.07.016
- Lenz N (2012) Von Schmetterlingen und Donnerdrachen. Natur und Kultur in Bhutan. Karlsruher Naturhefte 4, Naturkundemuseum Karlsruhe, 124 pp.
- Lindberg K (1932) Snakes of the Barsi Light Railway (Deccan). Journal of the Bombay Natural History Society 35(3): 690–697.
- Lindberg K (1939) Notes sur une Collection de Serpents du Deccan (Inde). Bulletin de la Société Zoologique de France 64: 328–336.
- Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, diferentiis, syonymis, locis. Editio decima, reformata. Tomus I. Laurentius Salvius, Holmiae [Stockholm], IV + 823 + (1) pp. [reprinted 1956, British Museum (Natural History), London].
- Lobo AS (2002) A preliminary checklist of snakes of the Bondla Wildlife Sancutuary, Goa. Cobra 49: 3–7.
- Madawala M, Surasinghe T, de Silva A, Gabadage D, Botejue M, Peabotuwage I, Kandambi D, Karunarathna S (2019) Reappraisal of herpetofauna recorded from Jaffna Peninsula in northern Sri Lanka with remarks on conservation, diversity, and distribution. Russian Journal of Herpetology 26(5): 247–260. https://doi.org/10.30906/1026-2296-2019-26-5-247-260
- Mahajan KK, Agrawal HP (1976) Report on a collection of reptile fauna from Simla Hills, Himachal Pradesh. Records of the Zoological Survey of India 71: 301–312.
- Mahendra BC (1984) Handbook of the snakes of India, Ceylon, Burma, Bangladesh, and Pakistan. The Annales of Zoology, Agra - 2 (India), 22, 412 pp.
- Manhas A, Kotwal A, Wanganeo RRN, Wanganeo A (2015a) Diversity, threats and conservation of herpetofauna in and around Barkatullah University, Bhopal (MP), India. International Journal of Advanced Research 3(9): 1546–1553.
- Manhas A, Raina R, Wanganeo A (2015b) Snakes of the Bhopal district, Madhya Pradesh, India with special reference to road mortality. Journal of Research in Biology 5(7): 1868–1873.
- Manhas A, Raina R, Wanganeo A (2018) Reptilian diversity of the Bhopal region in the state of Madhya Pradesh in Central India. IRCF Reptiles & Amphibians 25(2): 104–114.
- Marshall J (1832) A new universal gazetteer, containing a description of the principal nations, empires, kingdoms, states, provinces, cities, towns, forts, seas, harbours, rivers, lakes, canals, mountains, volcanoes, capes, caverns, cataracts and grottoes, of the known world, [etc., etc.] originally compiled by R. Brookes, M.D. W.W. Reed & Co., New York, 790+(24) pp.
- Martyn JH (1923) Martyn’s notes on Jaffna. Chronological, Historical, Biographical. American Ceylon Mission Press, Tellippalai, 363+xlvi+xx pp.
- Mason F (1861) Burmah, its people and natural productions or notes on the nations, fauna, flora, and minerals of Tenasserim, Pegu and Burmah, with systematic catalogues of the known mammals, birds, fish, reptiles, insects, mollusks, crustaceans, annalids, radiates, plants and minerals, with vernacular names. Trubner & Co., London, [1860], xvii+913 pp. [second edition]
- McClelland J, Griffith W (1844) Hodgson’s Illustrations of the Zoology of Nipal and Tibet. Calcutta Journal of Natural History and Miscellany of the Arts and Sciences 4: 129–130.
- Melvinselvan G, Nibedita D, Magesh P, Elambharathy K (2018) New locality records of Nagarjunsagar Racer, Platyceps bholanathi, Sharma, 1976 from Tamil Nadu, southern India. Captive & Field Herpetology 2(1): 29–32.
- Mertens R (1969) Die Amphibien und Reptilien West-Pakistans. Stuttgarter Beiträge zur Naturkunde 197: 1–96.
- Merrem B (1820) Versuch eines Systems der Amphibien/ Tentamen Systematis Amphibiorum. J. C. Krieger, Marburg, xv, xv+191, 191 pp., 1 pl. [bilingual German/Latin].
- Miller MA, Pfeiffer W, Schwartz T (2010) Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, Louisiana, 14 November 2010, pp. 1–8, https://doi.org/10.1109/GCE.2010.5676129
- Minton SA [jr] (1962) An annotated key to the amphibians and reptiles of Sind and Las Bela, West Pakistan. American Museum Novitates 2081: 1–60.
- Minton SA [jr] (1966) A contribution of the herpetology of West Pakistan. Bulletin of the American Museum of Natural History 134(2): 27–184, pl. 9–36.
- Minton SA [jr] (1990) Venomous bites by nonvenomous snakes: an annotated bibliography of colubrid envenomation. Journal of Wilderness Medicine 1: 119–127.
- Mirza ZA, Vyas R, Patel H, Maheta J, Sanap RV (2016) A new Miocene-divergent lineage of Old World Racer snake from India. PlosOne 11(3): 1–17. https://doi.org/10.1371/journal.pone.0154301
- Mohalik RK, Sahu SB, Arif M, Kar NB (2019) Bungarus caeruleus (Common Krait). Colouration and diet. Herpetological Review 50(1): 150–151.
- Müller F (1878) Katalog der im Museum und Universitätskabinett zu Basel aufgestellten Amphibien und Reptilien nebst Anmerkungen. Verhandelungen der naturforschenden Gesellschaft in Basel 6(4): 561–709, pl. 1–3.
- Müller F (1884) Dritter Nachtrag zum Katalog der herpetologischen Sammlung des Basler Museums. Verhandelungen der naturforschenden Gesellschaft in Basel 7(2): 274–299.
- Mukherjee D, Bhupathy S (2007) A new species of Wolf Snake (Serpentes: Colubridae: Lycodon) from Anaikatti Hills, Western Ghats, Tamil Nadu, India. Russian Journal of Herpetology 14(1): 21–26. https://doi.org/10.30906/1026-2296-2007-14-1-21-26
- Mullan JP (1927) A list of snakes and lizards from Panchgani. Journal of the Bombay Natural History Society 32(2): 380–381.
- Murthy TSN (1985) :Classification and distribution of the reptiles of India. The Snake 17: 48–71.
- Murthy TSN (1990) Illustrated guide to the Snakes of the Western Ghats, India. Records of the Zoological Survey of India, Kolkata, Occasional Papers 114, 76 pp.
- Murthy TSN (1992) An updated systematic index and bibliography of the reptiles of the Western Ghats, India. Records of the Zoological Survey of India 91(2): 169–180.
- Murthy TSN, Acharjyo LN (1987) Report on a collection of snakes in and around Nandankanan Biological Park, Orissa, India. The Snake 19: 139–142.
- Myers CW, Campbell JA (1981) A new genus and species of colubrid snake from the Sierra Madre del Sur of Guerrero, Mexico. American Museum Novitates 2708: 1–20.
- Negi S (1992) Himalayan Wildlife Habitat and Conservation. Indus Publ. Co., New Delhi, 207 pp.
- Nicholls L (1929) The identification of land snakes of Ceylon. Ceylon Journal of Science, Section D 2(3): 91–157.
- Nicholson E (1874) Indian Snakes. An elementary treatise on ophiology with a descriptive catalogue of the snakes found in India and the adjoining countries. Higginbotham & Co., Madras, 8+188 pp., 22 pls. [second edition].
- O’Shea M, Kusuma KI, Kaiser H (2018) First record of the Island Wolfsnake Lycodon capucinus (H. Boie in F. Boie 1827), from New Guinea, with comments on its widespread distribution and confused taxonomy, and a new record for the Common Sun Skink, Eutropis multifasciata (Kuhl 1820). IRCF Reptiles & Amphibians 25(1): 70–84.
- Pallas PS [1814] Zoographia Rosso-Asiatica, sistens omnium animalium in extenso Imperio Rossico et adjacentibus maribus observatorum recensionem, domicilia mores et descriptiones, anatomen atque icones plurimorum. Volumen tertium. Animalia monocardia seu frigidi sanguinis imperii RossoAsiatici. Officina Caesarea Academiae Scientiarum (1811) , vii+428+cxxv pp. [for publication date see Hemming and Noakes (1958)].
- Paliwal GT, Bhandarkar SV (2017) Faunistic diversity of serpents (Reptilia: Squamata) in Navegaon National Park of NNTR Maharashtra India with special reference to their conservation. Indo American Journal of Pharmaceutical Sciences 4(10): 3439–3444.
- Palot MJ (2015) A checklist of reptiles of Kerala, India. Journal of Threatened Taxa 7(13): 8010–8022. https://doi.org/10.11609/jott.2002.7.13.8010-8022
- Palot MJ, Radhakrishnan C (2003) Check-list of snakes of Kerala with their Malayalam vernacular names. Reptile Rap 5: 3–6.
- Palumbi SR (1996) The polymerase chain reaction. In: Hillis DM, Moritz C, Mable BK, editors. Molecular systematics. 2nd ed. Massachusetts: Sinauer Associates; p. 205–247.
- Palumbi SR, Martin AP, Romano SL, McMillan WO, Stice L, Grabowski G. (1991) The Simple Fool’s Guide to PCR ver. 2. Honolulu: University of Hawaii.
- Pande S, Mahabal A, Datar M, Sharma RM, Pati SK, Deshpande P, Bastwade DB, Padhye A, Kulkarni A, Joshi R, Pandit R, Mujumdar N, Mhaske P, Chhaya K, Patil R, Patwardhan A (2013) Biodiversity of the Parvati-Pachgaon Hills: A `Habitat Island’ in Pune Metropolis. Ela Journal 2(1): 23–41.
- Pasar V (2018) Snakes–The beautiful people. A field guide to Hyderabad Karnataka region. Partridge Publishing, Gurgaon, xi+56 pp.
- Pasar V, Paul R (2016) A preliminary survey of snake fauna in and around Chincholi Forest, Kalaburagi district Karnataka, India. International Journal of Scientific Research in Science, Engineering and Technology 2(5): 342–344.
- Patankar P (2011) Community structure of reptiles in selected habitats of Gujarat with major emphasis on the distribution and ecobiology of lizards. PhD Thesis, Department of Zoology, Faculty of Science, The Maharaja Sayajirao University of Baroda, iv+227+18 pp.
- Patel H, Vyas R (2019) Reptiles of Gujarat, India: Updated checklist, distribution, and conservation status. Herpetology Notes 125: 765–777.
- Patel H, Vyas R, Tank SK (2015) On the distribution, taxonomy, and natural history of the Indian Smooth Snake, Coronella brachyura (Günther, 1866). Amphibian & Reptile Conservation 9(2) [General Section]: 120–125.
- Patel H, Vyas R, Dudhatra B, Naik V, Chavda A, Chauhan D, Vaghashiya A, Vagadiya R, Vaghashiya P (2019a) Preliminary report on herpetofauna of Mount Girnar, Gujarat, India. – Journal of Animal Diversity 1(2): 9–35.
- Patel H, Vyas R, Ramani D (2019b) Argyrogena fasciolata (Banded Racer). Reproduction / Clutch size. Herpetological Review 50(1): 148–149.
- Patil PJ (2018) An ecological analysis of forest accounting. Environmental Analysis & Ecology Studies 4(2): 1–9.
- Phipson HM (1886) Catalogue of snakes in the society’s collection. Journal of the Bombay Natural History Society [1886] 1: 84–86.
- Porob PC, Kulkarni RS, Giri VS (2014) Biodiversity of Goa. A concise field guide. Pugmark Art Gallery, Kolhapur, 253 pp.
- Pradhan MS (2004) Reptilia. In: Director of the Zoological Survey of India (Ed.) Fauna of Pench National Park (Maharashtra). Zoological Survey of India, Conservation Area Series 20, Kolkata, 93–122.
- Pradhan MS (2008) Reptilia. In: Director of the Zoological Survey of India (Ed.) Fauna of Goa. Zoological Survey of India, State Fauna Series 16, Kolkata, 281–364.
- Pradhan MS, Talmale SS (2015) All about snakes. Myths and facts. Nature Books India, New Delhi, vi+366 pp.
- Pradhan S, Mishra D, Sahu KR (2014) An inventory and assessment of snake diversity of Gandhamardan Hills Range of western Orissa, India. International Journal of Pure and Applied Zoology 2(3): 241–245.
- Prakash S, Raziuddin M (2009) Report on the snake fauna of Hazaribag District, Jharkhand, India. Columban Journal of Life Sciences 10(1&2): 9–14.
- Prater S (1924) The snakes of the Bombay Island and Salsette. Journal of the Bombay Natural History Society 30(1): 151–176.
- Purkayastha J (2013) An amateur’s guide to reptiles of Assam. EBH Publishers, Guwahati, (9)+146 pp.
- Rameshwaran M (2008) A short note on the Banded Racer Argyrogena fasciolata (Shaw, 1802). Cobra 2(4): 22–23.
- Rajabizadeh M, Pyron RA, Nazarov R, Poyarkov NA, Adriaens D, Herrel A (2020) Additions to the phylogeny of colubrine snakes in Southwestern Asia, with description of a new genus and species (Serpentes: Colubridae: Colubrinae). PeerJ 8:e9016: 25pp. https://doi.org/10.7717/peerj.9016
- Rajendran MV (1986) Snakes of our land. Jaya Publications, Palayamkottai, (15)+1–135+1–88+1–112+1–4+(32)+89–112+1-32 pp. [in Tamil].
- Rao KT, Ghate HV, Sudhakar M, Javed SMM, Krishna SR (2005) Herpetofauna of Nallamalai Hills with eleven new records from the region including ten new records from Andhra Pradesh. Zoos’Print Journal 20(1): 1737–1740, suppl. i–xi.
- Rambaut A, Suchard MA, Xie D, Drummond AJ (2014) Tracer. Version 1.6. Available from: http://beast.bio.ed.ac.uk. Tracer (accessed 27 July 2014).
- Raut SR, Deshbhratar SM, Mahaley JA, Hile VK, Singh AJ, Metha GU (2014) Recent studies on the biodiversity of snakes in Palghar region, Thane, Maharashtra, India. Advances in Applied Science Research 5(2): 373–381.
- R Core Team (2017) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
- Reddy YA, Sadasivaiah B, Indira P, Pullaiah T (2013) Herpetofauna of Thummalapalle uranium mining area, Andhra Pradesh, India. International Journal of Biodiversity and Conservation 5(8): 515–522. https://doi.org/10.5897/IJBC2013.0546
- Rees A (1819) Coluber. The Cyclopaedia; or, Universal Dictionary of Arts, Sciences, and Literature, London 9, 19 pp. [not paginated].
- Ronquist F, Teslenko M, van der Mark P, Ayres D, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. https://doi.org/10.1093/sysbio/sys029
- Russell P (1796) An account of Indian serpents, collected on the Coast of Coromandel; containing descriptions and drawings of each species; together with experiments and remarks on their several poisons. George Nicol, London, vii+91 pp., 46 pls.
- Russell P (1803) Descriptions and figures of two hundred fishes; collected at Vizagapatam on the coast of Coromandel. Vol. II. G. and W. Nicol, London, 85+(4) pp., pl. ci–ccviii.
- Sagadevan J, Ganesh SR, Anandan N, Rajasingh R (2019) Recent records of the Banded Racer Argyrogena fasciolata (Shaw, 1802) (Reptilia: Squamata: Colubridae) from southern Coromandel Coast, peninsular India. Journal of Threatened Taxa 11(5): 13567–13572. https://doi.org/10.11609/jott.4446.11.5.13567-13572
- Sanyal DP, Sur S (1995) Reptilia. In: Director of the Zoological Survey of India (Ed.) Fauna of Kanha Tiger Reserve Madhya Pradesh. Zoological Survey of India, Fauna of Conservation Areas 7, Kolkata, 51–62.
- Satish (2008) Snakes of Coorg. A field guide to the snakes of Kodagu. Coorg Wildlife Society, Madikeri, xii+143+(1) pp. [bilingual English/Kannada].
- Savage JM (1997) On terminology for the description of the hemipenis of squamate reptiles. Herpetological Journal 7: 23–25. https://doi.org/10.1590/S1984-46702011000400005
- Sawant NS (2011) Ecology and conservation of pit vipers along the Western Ghats (Goa). PhD Thesis, Department of Zoology, Goa University, (12)+184+(2), (Appendix 1)+(4), (Appendix 2)+(21) pp.
- Sayeswara HA, Jeevan EN, Ashashree HM, Naik KL, Kumara SN (2015) A check list of snakes in and around Shivamogga city corporation, Karnataka, India. International Journal of Science and Nature 6(3): 495–500.
- Schätti B (1987) The phylogenetic significance of morphological characters in the Holarctic racers of the genus Coluber Linnaeus, 1758 (Reptilia, Serpentes). Amphibia-Reptilia 8: 401–418. https://doi.org/10.1163/156853887X00171
- Schätti B, Tillack F, Kucharzewski C (2014) Platyceps rhodorachis (Jan, 1863) – a study of the racer genus Platyceps Blyth, 1860 east of the Tigris (Reptilia: Squamata: Colubridae). Vertebrate Zoology 64(3): 297–405.
- Schlegel H (1827) Erpetologische Nachrichten. Isis von Oken 20(3): col. 281–294.
- Schlegel H (1837) Essai sur la Physionomie des Serpens. J. Kips, J.Hz. and W.P. Stockum, La Haye, Partie Générale: (3)+(1)+xxviii+251 pp., Partie Descriptive: (1)+606+xv+(1) pp., Atlas: (1)+(1)pp., 21 pls., 3 maps, 2 tables.
- Schleich HH (1993) Contribution to the Systematics and to the Bibliography of the Amphibians and Reptiles from Nepal. Journal of the Nepal Research Centre (JNRC) 9: 141–168.
- Schleich HH, Kästle W (2000) Umweltverständnis und Nationalparks. In: Fuhlrott-Museum & Forschungsinstitut (Ed.) Bhutan – Druk Yul: Land des Donnerdrachen. Fuhlrott-Museum & Forschungsinstitut, Wuppertal, 11–23.
- Sclater W (1891) List of Snakes in the Indian Museum. Baptist Mission Press, Calcutta, x+79 pp.
- Seetharamaraju M (2014) Taxonomy, diversity, distribution and ecology of snakes in Telangana. PhD Thesis, Osmania University, Hyderabad, (3)+xxxvi+347 pp.
- Shah KB (1995) Enumeration of habitat, geographical and altitudinal distribution of the Amphibians and Reptiles of Nepal. Biodiversity Profiles Project Publication 2, Kathmandu, 60 pp., Appendix 1–4, Maps 1–19.
- Shah KB (1997) Amphibians and reptiles used as food and medicine in Nepal. Natural History Museum, Tribhuvan University, Swayambhu, Kathmandu, Nepal, 30+(1) pp.
- Shah KB (1998a) Checklist of the herpetofauna of Nepal with English and vernacular names. NAHSON Bulletin 8(1-4): 26–30.
- Shah KB (1998b) Snakes of Nepal. In: Joshi DD, Chhetri MK (Eds) Proceedings of the Public Health & Medical Officers Training in Snakebite Management in Nepal. National Zoonoses & Food Hygiene Research Centre (NZFHRC), Kathmandu, 24–28.
- Shah KB, Tiwari S (2004) Herpetofauna of Nepal: A conservation companion. IUCN Nepal, Kathmandu, viii+237 pp.
- Shah KB, Shrestha J, Thapa C (2003) Snakebite management guideline. HMG, Ministry of Health, Dept. Health Serv. , Epidemiology & Disease Control Division, Kathmandu, 57 pp.
- Sharma BD (1999a) Snakes and human ecosystem. In: Sharma BD, Kumari T (Eds) Snakes in India: a source book. Asiatic Publishing House, Delhi, 40–47.
- Sharma BD (1999b) Fauna of Indian snakes. In: Sharma BD, Kumari T (Eds) Snakes in India: a source book. Asiatic Publishing House, Delhi, 87–108.
- Sharma BD (1999c) Scientific and vernacular names of some common snakes of India. In: Sharma BD, Kumari T (Eds) Snakes in India: a source book. Asiatic Publishing House, Delhi, 109–114,
- Sharma BD (2002a) Fauna of Indian snakes: A checklist. In: Sharma BD (Ed.) Indian poisonous snakes (An ecological and clinical study). Anmol Publ. Pvt. Ltd., New Delhi, 39–60.
- Sharma BD (2002b) Scientific and vernacular names of some common snakes of India. In: Sharma BD (Ed.) Indian poisonous snakes (An ecological and clinical study). Anmol Publ. Pvt. Ltd., New Delhi, 61–65.
- Sharma HB, Warman M (2011) Diversity, ecological structure and conservation of herpetofauna in Trans Yamunatic Region of Mathura. International Journal of Research in Engineering & Applied Sciences 1(4): 57–66.
- Sharma RC (1976) [A letter to the editor] Some observations on ecology and systematics of Coluber bholanathi, a new species of snake (Reptilia: Squamata: Colubridae) from India. Comparative Physiology and Ecology 1(3): 105–107.
- Sharma RC (1982) Studies on the pholidosis and variability in characters showing sexual dimorphism in various species of Indian reptiles. Records of the Zoological Survey of India, Calcutta, Occasional Papers 35: 1–153.
- Sharma RC (2000) Reptilia. In: Director of the Zoological Survey of India (Ed.) Fauna of Gujarat (Part-1) Vertebrates. Zoological Survey of India, State Fauna Series 8, Calcutta, 243–297.
- Sharma RC (2003) Handbook–Indian snakes. Zoological Survey of India, Kolkata, xx+292 pp., 48 pls.
- Sharma RC (2007) Fauna of India and adjacent countries. Reptilia. Volume-III (Serpentes). Zoological Survey of India, Kolkata, xxvi+410 pp.
- Sharma RC, Diksit VK (1976) Occurrence of Banded Racer, Coluber fasciolatus Shaw (Serpentes. Colubridae) in Rajasthan. Newsletter of the Zoological Survey of India 2(4): 171.
- Sharma S (2004) Observation on some snake species of Kedernath WLS, Garhwal Himalaya. Reptile Rap 6: 3.
- Shaw G (1802) General zoology or systematic natural history. III (II). Amphibia. G. Kearsley, London, vii+(2)+303 pp. [reprinted 1999 by the Society for the Study of Amphibians and Reptiles, Ithaca, New York].
- Sheikh AH (2017) Study of snake (Serpentes: Squamata) fauna of Dumna Nature Park, Jabalpur, India. International Journal of Zoology Studies 2(6): 54–57.
- Shrestha RL, Shah KB (1985) Nepalka sarpaharu [Snakes of Nepal]. Tribhuvan University, Natural History Museum, Kathmandu, 84 pp. [in Nepali].
- Shrestha TK (1998) Country report for Nepal. Herpetofauna of Nepal: Present status, distribution and conservation. In: de Silva A (Ed.) Proceedings of the International Conference on the Biology and Conservation of Amphibians and Reptiles of South Asia, Sri Lanka, August 1–5, 1996. Amphibia and Reptile Research Organization of Sri Lanka (ARROS), Peradeniya, 26–46.
- Shrestha TK (2001) Herpetology of Nepal. A field guide to amphibians and reptiles of Trans-Himalayan Region of Asia. B. Shrestha Publ., Kathmandu, 8+280 pp.
- Shrestha TK (2003) Wildlife of Nepal. A study of renewable resources of Nepal Himalayas. Mrs. Bimala Shrestha, Kathmandu, (13), map, 32 pls., 720 pp. [second edition].
- Siler CD, Oliveros CH, Santanen A, Brown RM (2013) Multilocus phylogeny reveals unexpected diversification patterns in Asian wolf snakes (genus Lycodon). Zoologica Scripta 42: 262–277.
- Silvestro D, Michalak I (2012) raxmlGUI: a graphical front-end for RAxML. Organisms Diversity and Evolution 12: 335–337. https://doi.org/10.1007/s13127-011-0056-0
- Singh KP, Ngullie MM (2010) Working plan for the forests of Dahanu Forest Division for the period 2010–2011 to 2019–2020. Government of Maharashtra, Mumbai, 218 pp.
- Singh L (1972) Evolution of karyotypes in snakes. Chromosoma 38: 185–236.
- Smith HM, David P (1999) George Shaw and the herpetology volume in his series, General Zoology. In: Shaw, G. (1802): General zoology or systematic natural history; Vol. III, Amphibia. Society for the Study of Amphibians and Reptiles, Ithaca, New York, [Faksimile], 5–96.
- Smith MA (1943) Fauna of British India, Ceylon and Burma, including the whole of the Indo-Chinese Sub-Region. Reptilia and Amphibia. Vol. III-Serpentes. Taylor and Francis Ltd., London, xii+583 pp. [reprinted 1961 by Survey of India Press, Dehra Dun and 1981 by Today and Tomorrow’s Printers & Publishers, New Delhi].
- Smith MA (1928) The status of some recently described genera and species of snakes. The Annals and Magazine of Natural History, ser. 10, 1(4): 494–497.
- Somaweera R (2004) Sri Lankan colubrid snakes. Sri Lankan Naturalist 6(3–4): 32–46.
- Somaweera R (2006) Sri Lankawe Serpayin [The snakes of Sri Lanka]. Wildlife Heritage Trust of Sri Lanka, Colombo, 8+297 pp. [in Singhalese].
- Srikanthan AN, Kumar GC, Urs AJ, Ganesh SR (2018) Appearances are deceptive: molecular phylogeny recovers the Scaly Gecko Hemidactylus scabriceps (Reptilia: Squamata: Gekkonidae) as a member of a scansorial and rupicolous clade. Journal of Threatened Taxa 10(9): 12147–12162. https://doi.org/10.11609/jott.3964.10.9.12147-12162
- Srinivasulu C, Das I (2008) The herpetofauna of Nallamala Hills, Eastern Ghats, India: An annotated checklist, with remarks on nomenclature, taxonomy, habitat use, adaptive types and biogeography. Asiatic Herpetological Research 11: 110–131.
- Srinivasulu C, Srinivasulu B, Rao CAN (2006) Reptilian fauna of Nagarjunasagar Srisailam Tiger Reserve, Andhra Pradesh. Records of the Indian Museum 106(3): 97–122.
- Stamatakis A (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22(21): 2688–2690. https://doi.org/10.1093/bioinformatics/btl446
- Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30(9): 1312–1313. https://doi.org/10.1093/bioinformatics/btu033
- Steindachner F (1867) Reise der Österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter den Befehlen des Commodore B. von Wüllersdorf-Urbair. Zoologischer Theil. Erster Band. Reptilien. Kaiserlich-Königliche Hof- und Staatsdruckerei, Wien, 98 pp., 3 pls.
- Stoliczka F (1871a) Notes on some Indian and Burmese ophidians (Abstract). Proceedings of the Asiatic Society of Bengal, September issue: 191–192.
- Stoliczka F (1871b) Notes on some Indian and Burmese ophidians. Journal of the Asiatic Society of Bengal 40(2): 421–445, pl. 25+26.
- Swan LW, Leviton AE (1962) The herpetology of Nepal: A history, checklist, and zoogeographical analysis of the herpetofauna. Proceedings of the California Academy of Sciences 4. Ser. , 32(6): 103–147.
- Tambre GN, Chavan SP (2016) Snake Species Diversity of Swami Ramanand Teerth Marathwada University, Nanded, Maharashtra State, India. International Journal of Current Research and Academic Review 4(6): 105–115.
- Taylor EH (1950) A brief review of Ceylonese snakes. University of Kansas Science Bulletin 33(14): 519–603.
- Thakur KN (2010) Impact of special economic zone (SEZ) on birds in Uran, Maharashtra. MSc Thesis, Department of Environmental Science Institute of Science, Mumbai, (8)+44 pp.
- Thakur S (2011) A note on snakes of Kanha National Park and surrounding areas. Reptile Rap 11: 2–5.
- Themido AA (1941) Répteis e Batráquios das Colónias Portuguesas (Catálogo das Colecções do Museu Zoológico de Coimbra). Memórias e Estudos do Museu Zoológico da Universidade de Coimbra Ser. 1, 119: 1–28.
- Theobald W (1868a) Catalogue of Reptiles in the Museum of the Asiatic Society of Bengal. Journal of the Asiatic Society of Bengal, Extra Number, vi+82 pp., Appendix i–iii.
- Theobald W (1868b) Catalogue of the reptiles of British Birma, embracing the provinces of Pegu, Martaban, and Tenasserim; with descriptions of new or little-known species. Journal of the Linnean Society of London. Zoology 10: 4–67.
- Theobald W (1876) Descriptive Catalogue of the Reptiles of British India. Thacker, Spink and Co., Calcutta, 1–238, i–xxxviii, Appendix i–viii, 6 pls.
- Theobald W (1880) Reptilian Fauna. In: Sparman HR (Ed.) The British Burma Gazetteer, Volume I. Government Press, Rangoon, 605–640.
- Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research 22: 4673–4680. https://doi.org/10.1093/nar/22.22.4673
- Tiedemann F, Häupl M (1980) Typenkatalog der Herpetologischen Sammlung. Teil II: Reptilia. Kataloge der wissenschaftlichen Sammlungen des Naturhistorischen Museums in Wien, 4 (Vertebrata 2). Naturhistorisches Museum Wien, 80 pp.
- Traill TS (Ed.) (1843) Essay on the physiognomy of Serpents. By H. Schlegel [etc.]. – Maclachlan, Steward and Co., Edinburgh, vii+254+(1) pp., 2 pls., 1 map. [English translation of the „Partie Générale“ of Schlegel (1837)].
- Uetz P, Freed P, Hošek J (Eds) (2019) The Reptile Database. http://www.reptile-database.org [last accessed 01 February 2021].
- Underwood G (1967) A contribution to the classification of snakes. Trustees of the British Museum (Natural History), x+179 pp.
- Upadhye MV, Puranik VV, Dabholkar P, Jadhav U (2012) Herpetofauna of the Vidyanagari campus of the University of Mumbai, Maharashtra. Reptile Rap 14: 15–20.
- Vibhute SM (2018) Serpentine diversity of Atpadi city and adjoining area, Tal. Atpadi, Dist. Sangli (MS) India. International Journal of Life Sciences, special issue A 10: 77–82.
- Vijayaraghavan B, Ganesh SR (2011) The illustrated book of South Indian snakes. The Chennai Snake Park Trust, Chennai, (7)+80+(3) pp.
- Vogel G, David P (2019) A new species of the Lycodon fasciatus complex from the Khorat Plateau, eastern Thailand (Reptiles, Squamata, Colubridae). Zootaxa 4577(3): 515–528. https://doi.org/10.11646/zootaxa.4577.3.6
- Vyas R (1987) A list of the snakes of the Bhavnagar District, Gujarat State. Journal of the Bombay Natural History Society 84(1): 227–231.
- Vyas R (1999) Gujarati vernacular names of some reptiles. Reptile Rap 1(1): 3–5.
- Vyas R (2000a) A review of threats and conservation of the reptiles of Gir forest, Gujarat, India. Cobra 39: 16–20.
- Vyas R (2000b) A review of reptile studies in Gujarat state. Zoo’s Print Journal 15(12): 386–390.
- Vyas R (2001a) Comments on ‘A synopsis of the reptiles of Gujarat, India’. Hamadryad [2000] 25(2): 203–207.
- Vyas R (2001b) Comments on `Reptilia’ a chapter in ‘Fauna of Gujarat’: a recent publication of ZSI, India. Cobra 46: 17–25.
- Vyas R (2001c) Reptile richness and diversity in and around the Gir Forest, Gujarat, India. Tigerpaper 28(3): 15–19.
- Vyas R (2004) Herpetofauna of Vansda National Park, Gujarat. Zoo’s Print Journal 19(6): 1512–1514.
- Vyas R (2007) Herpetofauna of Purna Wildlife Sanctuary, Gujarat, India. Raptile Rap 8: 10–15.
- Vyas R (2011) Reptilian diversity in and around the Shoolpaneshwar Wildlife Sanctuary, Gujarat, India. Reptile Rap 11: 5–15.
- Vyas R (2013) Snake diversity and voluntary rescue practice in the cities of Gujarat state: an evaluation. Reptile Rap 15: 27–39.
- Wagler J (1830) Natürliches System der Amphibien, mit vorangehender Classification der Säugethiere und Vögel. J. G. Cotta, München, Stuttgart und Tübingen, vi+354 pp., 1 pl., 1 table; Atlas: (4) pp, 7 pls.
- Wall F (1907) Notes on Snakes collected in Fyzabad. Journal of the Bombay Natural History Society 18(1): 101–129.
- Wall F (1908) The poisonous terrestrial snakes of our British Indian Dominions and how to recognize them. Bombay Natural History Society, viii+(1)+(1)+68+(1) pp. [second edition].
- Wall F (1913) The poisonous terrestrial snakes of our British Indian Dominions (including Ceylon) and how to recognize them, with symptoms of snake poisoning and treatment. Bombay Natural History Society, x+(2)+(1)+(1)+149+iv+(1) pp. [third edition].
- Wall F. (1914) A popular treatise on the common Indian snakes, Part xxii. Journal of the Bombay Natural History Society 23(1): 34–43, pl. 22.
- Wall F (1921) Ophidia Taprobanica or the snakes of Ceylon. H. R. Cottle, Government Printer, Colombo, 581 pp.
- Wall F (1924) Hand-list of the snakes of the Indian Empire. Part. II. Journal of the Bombay Natural History Society 29(3): 598–632.
- Wallach V, Williams KL, Boundy J (2014) Snakes of the World. A catalogue of living and extinct species. CRC Press, Taylor & Francis, Boca Raton, xxvii+1227 pp. https://doi.org/10.1201/b16901
- Walmiki NW, Awsare V, Karangutkar S, Wagh V, Yengal B, Salvi S, Pillai R (2012a) Herpetofauna of Maharashtra Nature Park, Mumbai, Maharashtra (India). World Journal of Environmental Biosciences 1(2): 90–99.
- Walmiki N, Karangutkar S, Yengal B, Kayande M, Wagh V, Pillai R, Dalvi S (2012b) Herpetofauna of Bassein Fort and surrounding region, Thane, Maharashtra, India. Trends in Life Sciences 1(3): 1–11.
- Waltner RC (1973) Geographical and altitudinal distribution of amphibians and reptiles in the Himalayas (Part 3). Cheetal 16(3): 14–19.
- Weinstein SA, Warrell DA, White J, Keyler DE (2011) „Venomous“ bites from non-venomous snakes. Elsevier, London, xxvii + 336 pp.
- Welch KRG (1983) A comment on the generic allocation of the Old World species of the genus Coluber. Litteratura Serpentium (English Edition) 3(4): 104–110.
- Welch KRG (1988) Snakes of the Orient: A checklist. R. E. Krieger Publ. Comp. Malabar, FL, vii+183 pp.
- Werner F (1924) Neue oder wenig bekannte Schlangen aus dem Naturhistorischen Staatsmuseum in Wien. Sitzungsberichte Österreichische Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse (Abt. 1), 133 (1-3): 29–56.
- Werner F (1929) Übersicht der Gattungen und Arten der Schlangen aus der Familie Colubridae. III. Teil (Colubrinae). Mit einem Nachtrag zu den übrigen Familien. Zoologische Jahrbücher (Systematik, Ökologie und Geographie) 57(1/2): 1–196.
- Whitaker R (1978) Common Indian snakes. A field guide. MacMillan India Ltd., Delhi, xiv+154 pp.
- Whitaker R (1982) Common Indian snakes. A field guide. MacMillan India Ltd., Delhi, xiv+154 pp. [reprint of 1978 edition].
- Whitaker R (2006) Common Indian snakes. A field guide. MacMillan India Ltd., New Delhi, xiv+138 pp. [sixth revised edition].
- Whitaker Z, Whitaker R (1986) The snakes around us. National Book Trust, India, New Delhi, 62 pp.
- Whitaker R, Captain A (2004) Snakes of India. The field guide. Draco Books, Chennai, xiv+480 pp. [abridged version in Tamil published in 2006].
- Wickham H (2017) Tidyverse: Easily Install and Load the ‘Tidyverse’. R package version 1.2.1.
- Wilkes J (1810) Encyclopaedia Londinensis; or, Universal Dictionary of Arts, Sciences, and Literature. Comprehending, under one general alphabetical arrangement, all the words and substance of every kind of dictionary extant in the English language. [etc.]. Compiled, digested, and arranged, by John Wilkes, of Milland House, in the county of Sussex, esquire; assisted by eminent scholars of the English Scotch, and Irish, universities, Vol. IV, (1)+(2)+911+(1) pp., 67 pls. [Coluber pls. 47–62].
- Willey A (1906) Terrestrial Colubridae of Ceylon. Spolia Zeylanica 3: 227–234.
- Williams KL, Wallach V (1989) Snakes of the world. Synopsis of snake generic Names. R. E. Krieger Publ. Comp. Malabar, FL, (1)+234 pp.
- Wilson LD (1967) Generic reallocation and review of Coluber fasciolatus Shaw (Serpentes: Colubridae). Herpetologica 23(4): 260–275.
- Wright GN (1834) A new and comprehensive gazetteer; being a delineation of the present state of the world, [etc., etc.], Vol. I. Thomas Kelly, London, clviii+592 pp.
- Zaher H (1999) Hemipenial morphology of the South American xenodontine snakes, with a proposal for a monophyletic Xenodontinae and a reappraisal of colubrid hemipenes. Bulletin of the American Museum of Natural History 240: 1–168.
- Zaher H, Murphy RW, Arredondo JC, Graboski R, Machado-Filho PR, Mahlow K, Montingelli GG, Botallo Quadros A, Orlov NL, Wilkinson M, Zhang Y-P, Grazziotin G (2019) Large-scale molecular phylogeny, morphology, divergence-time estimation, and the fossil record of advanced caenophidian snakes (Squamata: Serpentes). PloS ONE 14(5): 1–82. https://doi.org/10.1371/journal.pone.0216148
Appendices
Note 1
Francis Gowdie (date of birth unknown –12. Sept. 1813), served in the Third (1789–92) and Fourth (1798–99) Anglo-Mysore War and from 1805 to 1813 as Major General in the service of the East Indian Company Army at Madras (Chennai, Tamil Nadu state of India) (
Earlier attempts by
But “Caser cotta” lies on the northern part of the Malabar Coast and therefore opposite of the Coast of Coromandel where Russell spend most of the time during his medical service for the East Indian Company. Furthermore,
Note 2
The description of Coluber curvirostris published 1839 in Theodore E. Cantor’s “Spicilegium Serpentium Indicorum” is a very short bilingual (Latin/English) section of nine lines of text only. Cantor’s unpublished original manuscript entitled “Indian Serpents–Innocuous–Collected, figured & described (1831–1837)” is in the possession of the Bodleian Library, Oxford, U.K. This manuscript was the basis for
We here provide the transcript of the detailed description of C. curvirostris extracted from Cantor’s manuscript. The description is accompanied by the coloured plate no. 7 [reproduced here as Fig.
The title page contains following additional text: “A catalogue of the species will appear in the Proceedings of the Zool. Soc. London. April 29. 1839”, and the following information about the content of the manuscript: “35 Drawings {28 Figures of Serpents. New Species., 7. Anatomical details of Do.”, and “ The descriptions occupy 240 pages”.
The description of C. curvirostris occupies the manuscript pages 37 to 44 as follows:
“Coluber curvirostris/ Fig: 7./ C. supra partim late/olivaceo-viridis punctis/ & lineis obliquis al-/ bis nigrisquae, - par-/ tim aeneus; abdomine subfusco/ Scuta abdominalia …… 220./ Scutella subcaudalia 85./ Habitat: Bengal./
Above partly bright/ olive green with white/ and|<38>|black dots & oblique/ lines, - partly bron-/ ce-coloured; light/ yellow underneath./ Vernacular name:/ Tukkr-Bora./ The head a little broader,/ than the neck, subo-/ vate & strongly ar-/ ched, so that the/ profil of the rostrum/ is very oblique; the/ upper jaw extends/ about a line & a/ half beyond the low-/ er; |<39>| The nostrils moderate/ between two scuta./ The eyes, surrounded/ by two prae- & one post/ orbital scutum, large/ dark brilliant with a/ narrow reddish golden iris &/ a small round black/ pupil. The gape wide,/ straight with 8/8 labial/ scuta & with numerous little/ sharp teeth of equal size in the/ palaté & jaws. The/ tongue jet-black bi-/ furcate./ The shape of the body/ is elegant cylindrical;/ the tail more slender/ than |<40>| the trunk, gradually/ tapering into a small sharp point./ The scales of the body/ are oval-rhomboidal,/ very slightly imbricate,/ those of the tail shorter/ approaching a square/ form. The length of/ the abdominal scuta/ is about two lines;/ their breadth six lines./ The scutum of the/anus bipartite._/ The general colour/ is a shining bright/ olive green; on the an-/ terior part of the back,/ where the |<41>| sky-blue skin is visib-/ ble between the scales,/ a number of white & black dots, transversal-/ ly arranged, some of/ them forming rings./ About six inches from/ the head these dots change/ into white & bluish/ black oblique lines/ with points/ towards the head, ceasing/ on the posterior,/ third part of the body/ where the general color/ changes to a light/ brown |<42>| with a green metal-/ lic lustre./ The lips, the throat/ & the abdominal/ surface are of a light/ yellow./
Dimensions/
ft. in lin
Breadth of the head 0. 0. 5/
Length of the head* 0. 1. 0/
Length of the trunk 2. 4. 6/
Length of the tail 0. 7. 0/
Total length …… 3 ft. 0 in. 6 lin/ Circumference of the neck. 1 inch 3 lines./ Greatest circumf. of the trunk: 2 inches./
*The upper surface of the/ head being strongly arched,/ the measurement is taken on/ the lower surface from the projecting muzzle to the anterior margin/ of the third scutum abdominal |<43>| This not very common/ species is found in jungle although/ it may occasionally/ enter houses in/ search of mice & small/ lizards. It will sponta-/ neously starve for a/ considerable length of/ time: a female specimen/ in my possession lived/ upwards of eight months/ without food, during/ which period the skin/ was changed three times/ Having been shut up/ in a cage for more,/ than|<44>| five months, the cold/ weather inclusively, she/ brought forth in the commencement of May 1836 an egg/ of a cylindrical form, /1 ½ inch long, & half/ and inch in diameter/ of a light reddish/ yellow color. It was/ left in the cage for/ about a forthnight,/ until putrefaction required/ it removed. On opening/ the soft external mem-/ brane, it was found/ contain nothing but yolk./ Only when about changing/ the skin she became irritable /& attempted to bite as repre/ sented in the sketch./ This serpent is seldom/ found beyond three feet/ or so in length.”
Note 3
The alleged collector Sir William Crofton Twynam (1827–1922) also known as “Raja of the North” was born in Galle. He entered the Ceylon Civil Service in 1845 and held from 1869 until his retirement in 1896 the post as Government Agent on Jaffna (
Appendix 1
GenBank sequences number and locations for the samples used in this study. “_” no data or information not available. Sequences generated in this study marked in bold.
| Species | Locations | Voucher number | cytb | ND4 | CMOS | 16S | 12S | RAG1 | Genseq |
|---|---|---|---|---|---|---|---|---|---|
| Bamanophis dorri | Niokolo-Koba NP, Senegal | HLMD RA2906 | AY188040 | AY487042 | AY188001 | AY188081 | — | — | nomenclature |
| Bamanophis dorri | Niokolo-Koba NP, Senegal | HLMD RA2906 | AY188040 | AY487042 | AY188001 | AY188081 | — | — | |
| Dolichophis andreanus | Iran | ICSTZM.7H.1154 | MN531565 | MN531564 | — | — | AY647225 | — | |
| Dolichophis caspius | Greece, Serifos | NHMW KCC1 | AY376739 | AY487039 | AY376797 | AY376768 | AY039126 | — | |
| Dolichophis jugularis | Turkey, Antalya Province | MVZ 230242 | AY486917 | AY487046 | AY486941 | AY376769 | AY039152 | — | |
| Dolichophis schmidti | Turkmenistan |
|
AY486923 | AY487054 | AY486947 | AY376772 | AY039159 | — | |
| Eirenis aurolineatus | Turkey | CS4655/J170 | AY376749 | AY487070 | AY376807 | AY376778 | — | — | |
| Eirenis barani | Anavarza, Turkey & Osmaniye, Turkey | CS97 Eb1/J252 | AY376764 | — | AY376804 | AY376793 | — | — | |
| Eirenis collaris | Aras, Armenia | ZISP 27859 | AY376766 | — | AY376824 | AY376795 | — | — | |
| Eirenis coronelloides | Jordan | CS98 cr/J209 | AY376758 | — | AY376816 | AY376787 | — | — | |
| Eirenis decemlineata | Syria | CS94 de2/J226 | AY376760 | — | AY376818 | AY376789 | — | — | |
| Eirenis eiselti | Kahramanmaras, Turkey | CS4653/J166 | AY376747 | AY487069 | AY376805 | AY376776 | — | — | |
| Eirenis hakkariensis | Turkey | NMPGV 34464 | AY376761 | AY376819 | AY376790 | — | — | ||
| Eirenis levantinus | Antakya, Turkey & Osmaniye, Turkey | J168, CS4651/J171 | AY376765 | AY487071 | AY376806 | AY376794 | — | — | |
| Eirenis lineomaculata | Turkey | CS93li/J249 | AY376762 | — | AY376820 | AY376791 | — | — | |
| Eirenis medus | Hamadan, Iran | MNHG 2627.004 | AY376767 | — | AY376825 | AY376796 | AY647226 | — | |
| Eirenis modestus | Sivas, Turkey & W. Turkey | CS4664/J173 | AY376751 | AY487072 | AY376809 | AY376780 | AY039143 | — | |
| Eirenis persicus | Ufra, Halfeti, Turkey | CS00pe/J200 | AY376757 | — | AY376815 | AY376786 | AY647227 | — | |
| Eirenis punctatolineatus | Turkey | J175 | AY376755 | AY487073 | AY376810 | AY376784 | — | — | |
| Eirenis rothi | Turkey | CS97 ro1/J225 | AY376759 | — | AY376817 | AY376788 | — | — | |
| Eirenis thospitis | Van, Turkey | CS 01th/J194 | AY376754 | — | AY376812 | AY376783 | — | — | |
| Grayia ornata | Ogooue, Gaboon | — | — | — | AY611957 | AY611866 | AF158434 | — | |
| Hemerophis socotrae | Socotra, Yemen | HLMD RA2973 | AY188042 | AY487055 | AY188003 | AY188083 | AY039132 | — | |
| Hemorrhois algirus | Morocco | HLMD RA-1187 | AY486911 | AY487037 | AY486935 | AY611849 | AY039149 | — | |
| Hemorrhois hippocrepis | Cadiz Province, Spain & Morroco | MNCN 11988 | AY486916 | AY487045 | AY486940 | AY643350 | AY039139 | KY762199 | |
| Hemorrhois nummifer | Kotajsk Region, Armenia | ZISP 27709 | AY376742 | AY487049 | AY376800 | AY376771 | AY039163 | — | |
| Hemorrhois ravergieri | Kotajsk Region, Armenia | ZISP 27733 | AY486920 | AY487050 | AY486944 | — | AY039153 | — | |
| Hierophis cypriensis | Cyprus | Hcyp43 | JX315475 | — | — | — | AY541502 | — | |
| Hierophis gemonensis | Narina, Croatia | — | AY376741 | AY487044 | AY376799 | AY376770 | AY039145 | — | |
| Hierophis viridiflavus | Spain | MVZ 178418 | AY486925 | AY487057 | AY486949 | LN552128 | AY541507 | KY762200 | |
| Lytorhynchus diadema | Djébil NP, Tunisia | HLMD RA2333 | AY188025 | — | AY187986 | AY188064 | AY647229 | — | |
| Lytorhynchus gaddi | Khuzestan Province, Iran | MVZ234500 | KX909327 | — | — | — | KX909262 | — | |
| Lytorhynchus maynardi | Pakistan | MVZ248462 | KX909344 | — | KX909367 | KX909317 | KX909285 | — | |
| Lytorhynchus paradoxus | Sam, Rajasthan, India | — | — | MZ020427 | — | MW991416 | — | — | genseq4- ND4, 16S |
| Macroprotodon abubakeri | Morocco | E2208.8 | AY643378 | — | — | AY643336 | AY643297 | — | |
| Macroprotodon brevis | Portugal | BMB990094 | KY762176 | — | KY762079 | KY762061 | KY762034 | KY762201 | |
| Macroprotodon brevis brevis | Te-touan, Tleta Tarhremt, Morocco | MVZ 186239 | DQ907242 | DQ902333 | — | — | — | — | |
| Macroprotodon cucullatus | Bou Hedma, Tunisia | HLMD RA2974 | AY188026 | — | AY187987 | AY188065 | AY643288 | — | |
| Macroprotodon cucullatus | Spain | MVZ 186073 | DQ902150 | AY487064 | AF471145 | — | — | — | |
| Macroprotodon mauritanicus | Mallorca, Baleanc I., Spain | E2208.6 | AY643384 | — | — | AY643342 | AY643301 | — | |
| Mopanveldophis zebrinus | Namibia |
|
AY188043 | AY487058 | AY188004 | AY188084 | — | — | |
| Muhtarophis barani | Turkey | ZMHRU2014/60-5 | KX909323 | — | KX909374 | KX909292 | KX909253 | — | |
| Orientocoluber spinalis | Yunnan, China | MVZ 211019 | AY486924 | AY487056 | AY376802 | AY376773 | AY541508 | — | |
| Persiophis fahimii | Iran | RAP-2019 | MN531567 | MN531566 | MT163748 | — | MN536809 | — | |
| Platyceps atayevi | Turkmenistan |
|
AY486912 | AY487038 | AY486936 | — | AY647231 | — | |
| Platyceps bholanathi | Hyderabad, India | — | — | MZ020435 | — | — | — | — | genseq4- ND4 |
| Platyceps collaris | Jordan | HLMD J14 | AY486922 | AY487053 | AY486946 | — | AY039133 | _ | |
| Platyceps plinii | Bangalore, India | NCBS AQ-492 | MZ020421 | MZ020428 | MZ020440 | MW991417 | — | — | genseq4- cytb, ND4, CMOS, 16S |
| Platyceps plinii | Baripada, India | ZSI-CZRC-V-6384 | — | MZ020432 | — | — | MK329299 | — | genseq4- ND4, 12S |
| Platyceps plinii | Choudwar, India | ZSI-CZRC-V-6329 | — | MZ020429 | MZ020441 | MW991418 | — | — | genseq4- ND4, CMOS, 16S |
| Platyceps plinii | Yavatmal, India | ZSI-CZRC-V-6413 | — | MZ020430 | — | — | — | — | genseq4- ND4 |
| Platyceps plinii | Pune, India | ZSI-CZRC-V-6416 | — | MZ020431 | — | — | — | — | geneseq4- ND4 |
| Platyceps florulentus | Egypt | — | AY486915 | AY487043 | AY486939 | — | AY039161 | — | |
| Platyceps gracilis | Gujarat, India | — | MZ020424 | MZ020436 | MZ020443 | — | — | — | genseq4- CTYB, ND4, CMOS |
| Platyceps josephi sp. nov. | Anaikatti, India | ZSI-CZRC-7358 | — | MZ020433 | — | MW991419 | — | — | genseq3- ND4, 16S |
| Platyceps josephi sp. nov. | Tuticorin, India | NCBS-AU732 | MZ020422 | MZ020434 | MZ020442 | — | — | — | genseq1- cytb, ND4, CMOS |
| Platyceps josephi sp. nov. | Karur, India | ZSI-CZRC-6639 | MZ020423 | — | — | MW991420 | — | MZ020447 | genseq2- cytb, 16S, RAG1 |
| Platyceps karelini | Krasnovodsk, Turkmenistan & Uzbekistan |
|
AY486918 | AY487047 | AY486942 | AY611820 | AY647232 | — | |
| Platyceps rhodorachis | Ahal Prov., Turkmenistan & Ambouli, Djibouti |
|
AY486921 | AY487051 | AY486945 | — | — | — | |
| Platyceps rogersi | Sinai, Egypt | NMW KCR2 | AY188041 | AY487052 | AY188002 | AY188082 | MF767306 | — | |
| Platyceps saharicus | Hurkania, Israel | SMNH<ISR>:17853 | MF767406 | — | — | — | MF767311 | — | |
| Platyceps ventromaculatus | Sam, Rajasthan, India | — | — | MZ020437 | MZ020444 | — | — | — | genseq4- ND4, CMOS |
| Rhynchocalamus arabicus | Oman | CN4780 | KT878854 | — | KT878851 | KT878847 | KT878842 | — | |
| Rhynchocalamus dayanae | Israel | TAU.R17093 | KX909332 | — | KX909361 | KX909313 | KX909283 | — | |
| Rhynchocalamus ilamensis | Ilam, Abdanan, Iran | — | KY777450 | — | KY777447 | — | — | — | |
| Rhynchocalamus melanocephalus | Israel | TAU.R16340 | KX909342 | — | KX909350 | KX909306 | KX909264 | — | |
| Rhynchocalamus satunini | Iran | CAS228723 | KT878855 | — | KT878850 | KT878846 | KT878843 | — | |
| Rhynchocalamus satunini | Turkey | ZMHRU2015/0 | KX909328 | — | KX909362 | KX909297 | KX909263 | — | |
| Scaphiophis albopunctatus | Tanzania | CMRK284 | DQ486345 | DQ486321 | DQ486169 | — | — | — | |
| Sibynophis subpunctatus | Sri Lanka | RAP0491 | KC347471 | KC347516 | KC347411 | KC347373 | KC347335 | — | |
| Spalerosophis diadema | Egypt | — | AF471049 | — | AF471155 | — | KC347335 | — | |
| Spalerosophis diadema | Jordan & Hail, Saudi Arabia | HLMD J62 | AY486926 | AY487059 | AY486950 | HQ658450 | AY039148 | — | |
| Wallaceophis gujaratensis | Gujarat, India | NCBS HA-105 | — | KR819923 | KR819917 | KR819921 | KR819920 | — | |
| Wallaceophis gujaratensis | Gujarat, India | — | MZ020425 | MZ020438 | MZ020445 | MW991421 | — | MZ020448 | genseq4- cytb, ND4, CMOS, 16S, RAG1 |
| Wallophis brachyura | Unknown | — | KX782330 | KX782331 | KX768756 | — | — | — | |
| Wallophis brachyura | Maharashtra, India | — | MZ020426 | MZ020439 | MZ020446 | MW991422 | — | MZ020449 | genseq4- cytb, ND4, CMOS, 16S, RAG2 |
Appendix 2
Partitions and models of sequence evolution used in the Maximum Likelihood (ML) and Bayesian Inference (BI) phylogenetic analyses. In the second column, 1st, 2nd and 3rd refer to codon position.
| Partitions | Sites | ML | BI |
| P1 | 12S, cytb 1st, ND4 1st | GTR+G | GTR+I+G |
| P2 | ND4 2nd , cytb 2nd | GTR+G | HKY+I+G |
| P3 | cytb 3rd ,ND4 3rd | GTR+G | GTR+I+G |
| P4 | cmos 1st, cmos 2nd, rag11st, rag1 2nd | GTR+G | HKY+G |
| P5 | cmos 3rd, rag1 3rd | GTR+G | HKY+G |
| P6 | 16S | GTR+G | GTR+I+G |
Appendix 3
Pairwise uncorrected genetic distance matrices showing within and between generic distances for the three mitochondrial genes. Within genus pairwise distances, Argyrogena – Platyceps and Wallaceophis – Wallophis genetic distances are marked in bold.
| cytb | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
| Argyrogena | 0.04–0.05 | ||||||||||||||||
| Bamanophis | 0.18–0.19 | ||||||||||||||||
| Dolichophis | 0.14–0.18 | 0.16–0.18 | 0.07–0.13 | ||||||||||||||
| Eirenis | 0.15–0.18 | 0.16–0.17 | 0.12–0.16 | 0.04–0.15 | |||||||||||||
| Hemerophis | 0.18 | 0.17 | 0.14–0.17 | 0.14–0.17 | |||||||||||||
| Hemorrhois | 0.14–0.17 | 0.15–0.17 | 0.12–0.17 | 0.13–0.17 | 0.16–0.18 | 0.10–0.13 | |||||||||||
| Hierophis | 0.15–0.17 | 0.17–0.18 | 0.12–0.15 | 0.10–0.15 | 0.15–0.16 | 0.13–0.15 | 0.09–0.10 | ||||||||||
| Lytorhynchus | 0.16–0.19 | 0.16–0.18 | 0.15–0.17 | 0.15–0.18 | 0.17 | 0.15–0.17 | 0.15–0.16 | 0.11–0.17 | |||||||||
| Macroprotodon | 0.17–0.19 | 0.12–0.15 | 0.13–0.18 | 0.14–0.19 | 0.15–0.17 | 0.15–0.19 | 0.15–0.18 | 0.14–0.18 | 0.01–0.14 | ||||||||
| Mopanveldophis | 0.19–0.21 | 0.20 | 0.18–0.19 | 0.17–0.20 | 0.19 | 0.18–0.19 | 0.19–0.20 | 0.18–0.20 | 0.15–0.18 | ||||||||
| Muhtarophis | 0.17–0.19 | 0.18 | 0.15–0.19 | 0.16–0.19 | 0.19 | 0.17–0.18 | 0.17–0.18 | 0.17–0.20 | 0.17–0.21 | 0.21 | |||||||
| Orientocoluber | 0.17 | 0.16 | 0.13–0.16 | 0.12–0.15 | 0.15 | 0.14–0.16 | 0.12–0.13 | 0.16–0.18 | 0.15–0.17 | 0.18 | 0.18 | ||||||
| Persiophis | 0.16–0.17 | 0.16 | 0.14–0.16 | 0.14–0.16 | 0.16 | 0.15–0.16 | 0.14–0.17 | 0.14–0.16 | 0.15–0.17 | 0.18 | 0.17 | 0.15 | |||||
| Platyceps | 0.13–0.17 | 0.17–0.18 | 0.13–0.18 | 0.13–0.18 | 0.16–0.17 | 0.13–0.16 | 0.14–0.17 | 0.15–0.19 | 0.14–0.20 | 0.18–0.20 | 0.17–0.19 | 0.15–0.16 | 0.15–0.17 | 0.08–0.15 | |||
| Rhynchocalamus | 0.17–0.19 | 0.16–0.18 | 0.15–0.18 | 0.15–0.19 | 0.16–0.17 | 0.15–0.18 | 0.15–0.18 | 0.15–0.17 | 0.14–0.19 | 0.20–0.21 | 0.18–0.20 | 0.16–0.17 | 0.14–0.16 | 0.15–0.20 | 0.03–0.13 | ||
| Scaphiophis | 0.19–0.21 | 0.19 | 0.17–0.20 | 0.18–0.20 | 0.20 | 0.18–0.20 | 0.19–0.20 | 0.18–0.19 | 0.18–0.20 | 0.20 | 0.20 | 0.20 | 0.18 | 0.19–0.22 | 0.19–0.20 | ||
| Spalerosophis | 0.15–0.17 | 0.16 | 0.15–0.16 | 0.13–0.16 | 0.16–0.17 | 0.14–0.15 | 0.14–0.16 | 0.16–0.18 | 0.14–0.18 | 0.18 | 0.18 | 0.16 | 0.15–0.16 | 0.14–0.16 | 0.17–0.19 | 0.19 | |
| Wallaceophis | 0.18–0.20 | 0.19 | 0.16–0.20 | 0.18–0.21 | 0.19–0.20 | 0.17–0.19 | 0.18–0.19 | 0.17–0.19 | 0.19–0.20 | 0.21–0.22 | 0.20–0.22 | 0.18–0.19 | 0.17 | 0.17–0.19 | 0.16–0.18 | 0.19–0.21 | 0.19 |
| Wallophis | 0.15–0.19 | 0.16–0.17 | 0.14–0.16 | 0.14–0.18 | 0.18–0.19 | 0.15–0.18 | 0.13–0.16 | 0.15–0.16 | 0.16–0.18 | 0.18–0.20 | 0.18–0.19 | 0.15–0.17 | 0.16 | 0.14–0.18 | 0.15–0.18 | 0.18–0.19 | 0.16 |
| ND4 | |||||||||||||||||
| Genus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
| Argyrogena | 0.05 | ||||||||||||||||
| Bamanophis | 0.14–0.15 | ||||||||||||||||
| Dolichophis | 0.16–0.18 | 0.13–0.15 | 0.05–0.13 | ||||||||||||||
| Eirenis | 0.14–0.19 | 0.14–0.15 | 0.12–0.16 | 0.07–0.11 | |||||||||||||
| Hemerophis | 0.16–0.17 | 0.14 | 0.14–0.16 | 0.16 | |||||||||||||
| Hemorrhois | 0.14–0.18 | 0.14–0.16 | 0.13–0.17 | 0.14–0.17 | 0.16–0.17 | 0.1–0.14 | |||||||||||
| Hierophis | 0.16–0.17 | 0.13–0.15 | 0.12–0.15 | 0.12–0.15 | 0.16–0.17 | 0.13–0.16 | 0.09 | ||||||||||
| Lytorhynchus | 0.20 | 0.18 | 0.17–0.19 | 0.17–0.19 | 0.18 | 0.17–0.19 | 0.17–0.18 | ||||||||||
| Macroprotodon | 0.15–0.16 | 0.13 | 0.13–0.15 | 0.13–0.15 | 0.15 | 0.12–0.15 | 0.13–0.14 | 0.16 | |||||||||
| Mopanveldophis | 0.18–0.19 | 0.16 | 0.16–0.17 | 0.16–0.18 | 0.18 | 0.17–0.18 | 0.15–0.16 | 0.17 | 0.15 | ||||||||
| Orientocoluber | 0.14–0.16 | 0.14 | 0.14–0.15 | 0.11–0.13 | 0.16 | 0.14–0.16 | 0.13–0.14 | 0.17 | 0.12 | 0.17 | |||||||
| Persiophis | 0.15–0.17 | 0.16 | 0.15–0.16 | 0.16–0.19 | 0.19 | 0.16–0.18 | 0.15–0.17 | 0.15 | 0.14 | 0.18 | 0.16 | ||||||
| Platyceps | 0.13–0.17 | 0.15–0.16 | 0.14–0.18 | 0.14–0.18 | 0.16–0.18 | 0.13–0.18 | 0.14–0.18 | 0.18–0.2 | 0.13–0.17 | 0.17–0.19 | 0.15–0.17 | 0.17–0.21 | 0.1–0.17 | ||||
| Scaphiophis | 0.20–0.21 | 0.18 | 0.18–0.19 | 0.20–0.21 | 0.18 | 0.18–0.20 | 0.18 | 0.20 | 0.18–0.19 | 0.19 | 0.19 | 0.20 | 0.19–0.22 | ||||
| Spalerosophis | 0.15 | 0.14 | 0.14–0.17 | 0.13–0.15 | 0.15 | 0.09–0.14 | 0.13–0.14 | 0.15 | 0.11 | 0.16 | 0.13 | 0.16 | 0.13–0.16 | 0.16 | |||
| Wallaceophis | 0.2–0.21 | 0.19 | 0.18–0.20 | 0.17–0.20 | 0.19 | 0.18–0.20 | 0.19 | 0.18–0.19 | 0.18 | 0.2–0.21 | 0.19 | 0.18 | 0.19–0.2 | 0.21 | 0.19 | ||
| Wallophis | 0.16–0.18 | 0.17 | 0.16–0.19 | 0.16–0.19 | 0.17–0.18 | 0.15–0.19 | 0.17–0.18 | 0.16–0.17 | 0.16 | 0.19–0.21 | 0.17 | 0.14–0.16 | 0.16–0.2 | 0.19–0.2 | 0.15–0.16 | 0.15–0.17 | |
| 16S | |||||||||||||||||
| Genus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
| Argyrogena | 0.03 | ||||||||||||||||
| Bamanophis | 0.08–0.1 | ||||||||||||||||
| Dolichophis | 0.08–0.1 | 0.06–0.07 | 0.02–0.04 | ||||||||||||||
| Eirenis | 0.06–0.11 | 0.06–0.08 | 0.03–0.06 | 0–0.06 | |||||||||||||
| Hemerophis | 0.09–0.11 | 0.08 | 0.06–0.07 | 0.07–0.08 | |||||||||||||
| Hemorrhois | 0.07–0.11 | 0.07–0.08 | 0.04–0.06 | 0.05–0.09 | 0.06–0.07 | 0.04–0.05 | |||||||||||
| Hierophis | 0.07–0.08 | 0.06–0.07 | 0.04–0.05 | 0.04–0.06 | 0.06 | 0.06–0.08 | 0.02 | ||||||||||
| Lytorhynchus | 0.08–0.11 | 0.07–0.1 | 0.06–0.08 | 0.05–0.08 | 0.07–0.09 | 0.06–0.09 | 0.06–0.08 | 0.04–0.07 | |||||||||
| Macroprotodon | 0.07–0.11 | 0.05–0.07 | 0.03–0.06 | 0.04–0.07 | 0.04–0.08 | 0.05–0.07 | 0.04–0.07 | 0.04–0.09 | 0.01–0.04 | ||||||||
| Mopanveldophis | 0.08–0.1 | 0.05 | 0.06–0.07 | 0.05–0.07 | 0.08 | 0.07 | 0.06 | 0.07–0.09 | 0.05–0.07 | ||||||||
| Muhtarophis | 0.07–0.08 | 0.07 | 0.05 | 0.05–0.07 | 0.08 | 0.06–0.07 | 0.05–0.06 | 0.05–0.07 | 0.05–0.07 | 0.07 | |||||||
| Orientocoluber | 0.06–0.07 | 0.06 | 0.04–0.05 | 0.03–0.06 | 0.06 | 0.05–0.06 | 0.04 | 0.05–0.07 | 0.04–0.06 | 0.06 | 0.05 | ||||||
| Platyceps | 0.05–0.07 | 0.07–0.08 | 0.06–0.07 | 0.05–0.08 | 0.07–0.08 | 0.05–0.07 | 0.06–0.07 | 0.06–0.08 | 0.06–0.08 | 0.07–0.08 | 0.05–0.06 | 0.05–0.06 | 0.04 | ||||
| Rhynchocalamus | 0.07–0.1 | 0.08 | 0.06–0.08 | 0.06–0.08 | 0.08–0.09 | 0.06–0.09 | 0.06–0.08 | 0.04–0.07 | 0.05–0.08 | 0.07–0.08 | 0.05–0.06 | 0.06–0.07 | 0.06–0.08 | 0–0.05 | |||
| Spalerosophis | 0.09–0.1 | 0.09 | 0.08 | 0.07–0.1 | 0.08 | 0.07–0.08 | 0.08 | 0.08–0.1 | 0.06–0.08 | 0.1 | 0.08 | 0.07 | 0.07–0.08 | 0.08–0.09 | |||
| Wallaceophis | 0.1–0.11 | 0.09 | 0.07–0.09 | 0.07–0.09 | 0.08 | 0.08 | 0.08 | 0.08–0.09 | 0.06–0.08 | 0.08 | 0.07–0.08 | 0.08 | 0.08 | 0.07–0.09 | 0.08–0.09 | ||
| Wallophis | 0.09–0.11 | 0.07 | 0.08–0.09 | 0.07–0.09 | 0.09 | 0.07–0.09 | 0.08 | 0.07–0.08 | 0.06–0.09 | 0.08 | 0.08 | 0.08 | 0.08–0.09 | 0.06–0.08 | 0.09 | 0.05 |
Appendix 4
BI phylogeny showing inferred phylogenetic relationships of old world racers. Labelled in pink color are the sequences generated in this study, colored blue is the sample from this study+GenBank sequence. Posterior probability support values shown at internal branches, values below 70 are not shown.
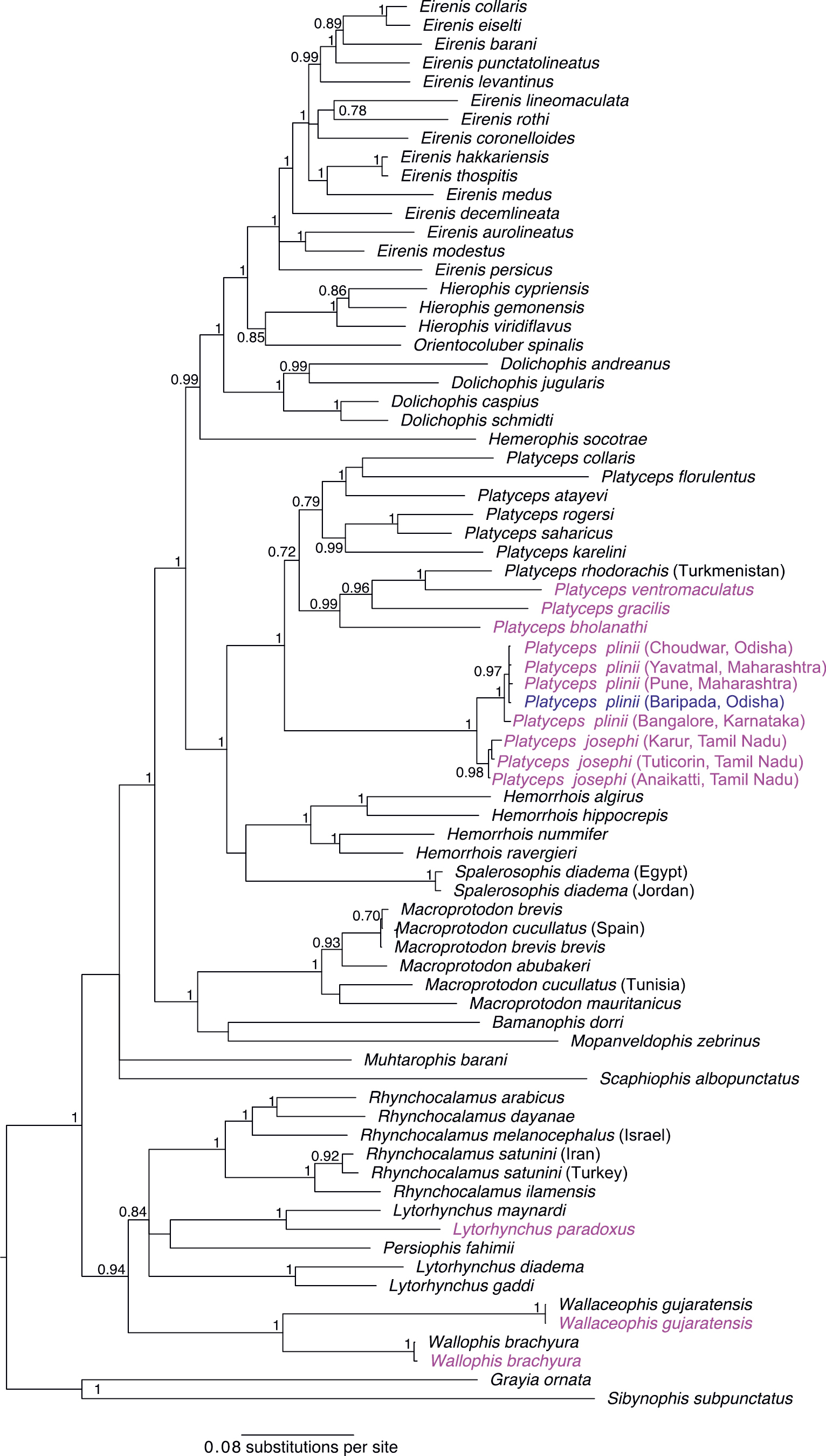
Appendix 5
Maximum Likelihood phylogeny constructed using cmos dataset. Number at internal branches are ML bootstrap support, values below 60 are not shown.
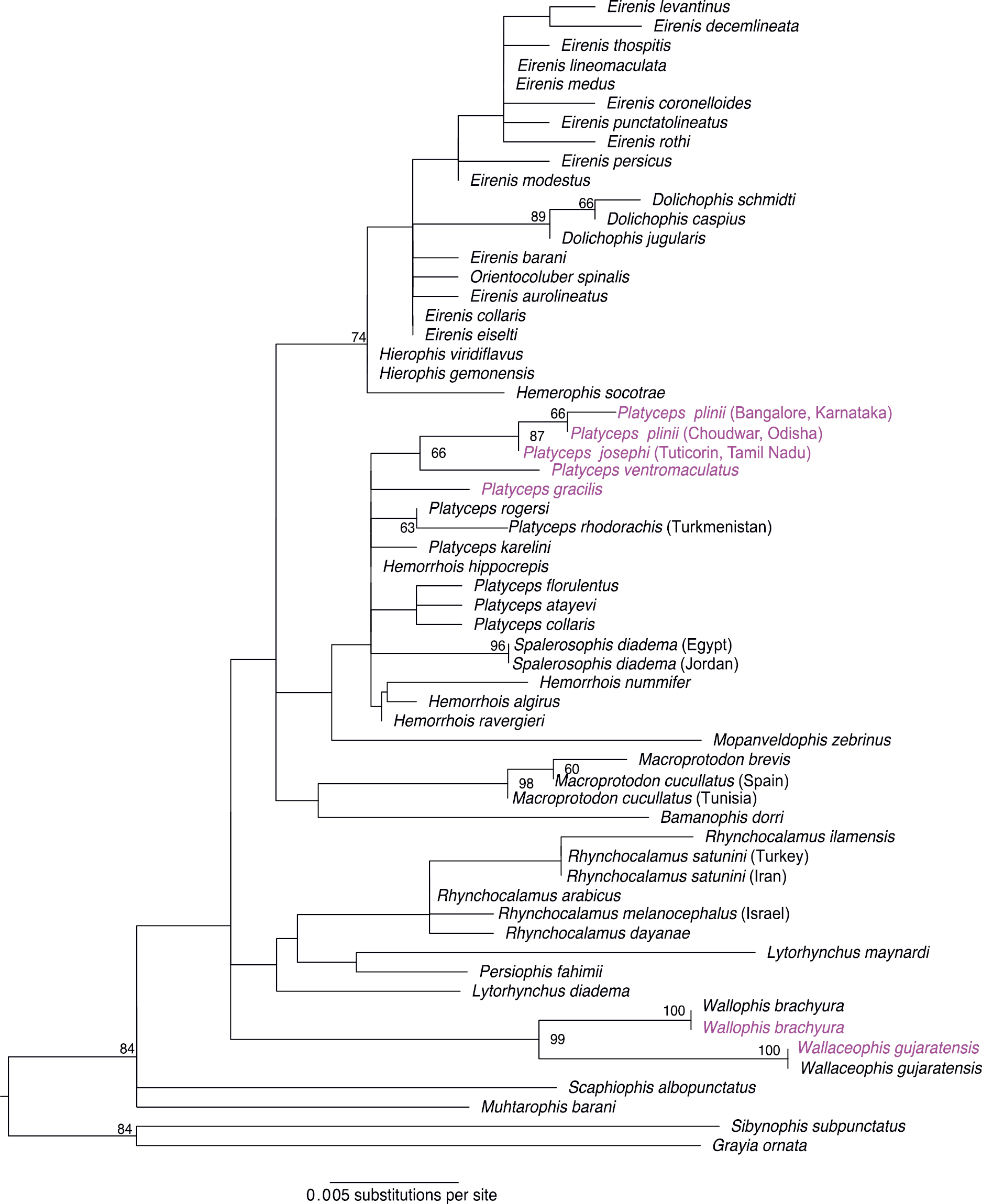
Appendix 6
Platyceps plinii comb. nov. (
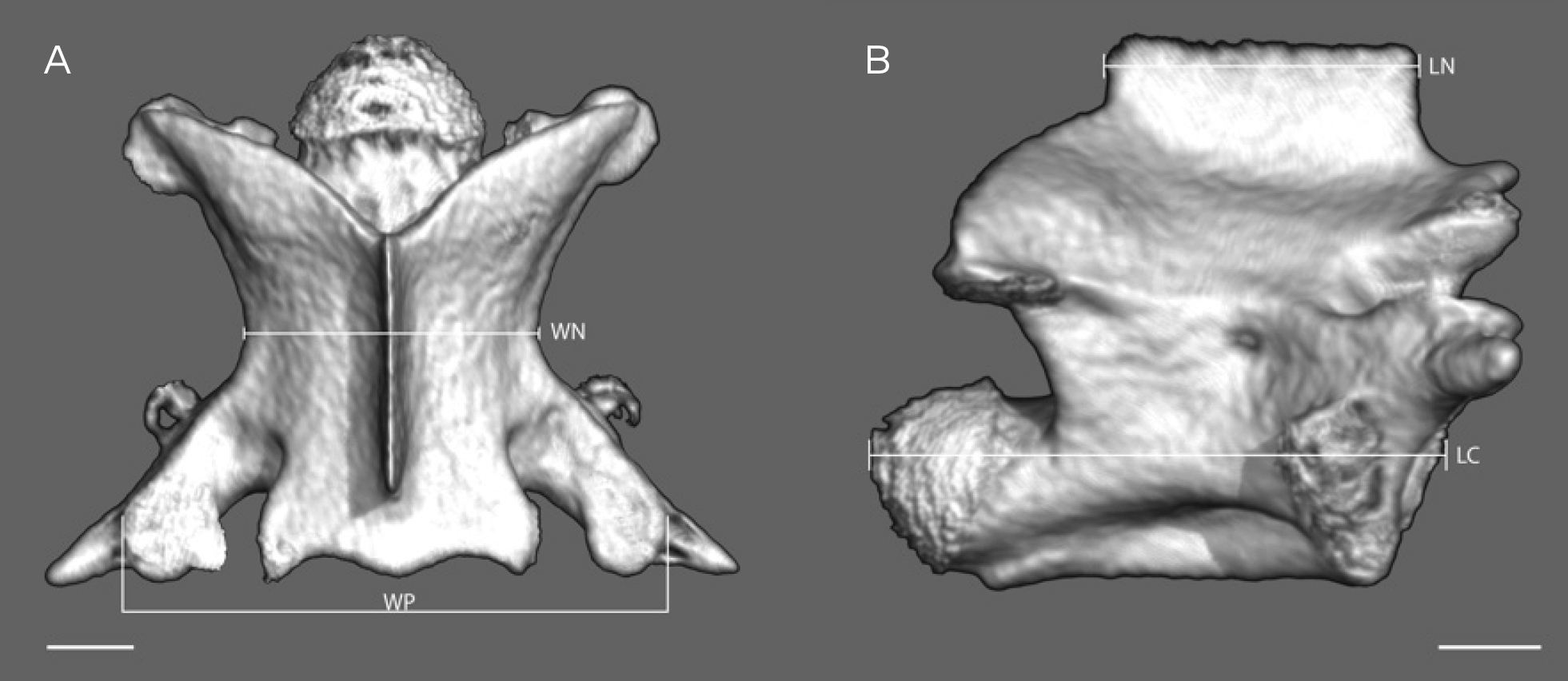
Appendix 7
Table with summarized characters used for osteological comparison of Platyceps spp. and related genera (Hemorrhois and Spalerosophis). LC/WN: ratio length of centrum to least width of neural arch, LC/WP: ratio length of centrum to distance between outer edges of articular facets of prezygapophyses, LN/WN: ratio length of neural spine to least width of neural arch. (1) Data from
| Taxon | Lateral processes of premaxilla | Ascending process of premaxilla | TLn/TWn | TLln/Lns | Mesial process of pterygoid | Pterygoid process at basioccipital | LC/WN | LC/WP | LN/WN |
| Platyceps plinii comb. nov. | individually short or long, one tip | short, in contact with nasals | 0.93–1.04 | 40%–50% | present only in 60% of examined specimens | individually unstructured or structured | 1.21–1.59 | 0.73–0.95 | 0.81–1.01 |
| Platyceps josephi sp. nov. | short, one tip | short, in contact with nasals | 1.16–1.18 | 47%–58% | present | individually unstructured or structured | 1.54 3 | 0.853 | 1.023 |
| Platyceps florulentus 1 | long, two tips | short, in contact with nasals | 1.24 | 68% | absent | structured | 1.70 [1.32–1.48] | 0.85 [0.68–0.79] | 1.15 [0.95–1.05] |
| Platyceps rhodorachis 2 | long, one tip or stout | long, not in contact with nasals | 1.27–1.39 | 60%–77% | absent | structured | 1.60–1.70 [1.41–1.55] | 0.83–0.88 [0.55–0.64] | 1.20–1.27 [1.09–1.25] |
| Platyceps ventromaculatus 1 | long, stout | long, in contact with nasals | 1.28 | 56% | absent | unstructured | 1.68 [1.49–1.51] | 0.86 [0.76–0.78] | 1.10 [1.16–1.18] |
| Hemorrhois nummifer 1 | / | / | / | / | / | / | [1.22–1.39] | [0.63–0.71] | [0.84–1.07] |
| Hemorrhois ravergieri 1 | / | / | / | / | / | / | 1.61 [1.20–1.41] | 0.86 [0.66–0.70] | 0.80 [0.87–1.16] |
| Spalerosophis diadema | / | / | / | / | / | / | 1.08 | 0.62 | 0.80 |
Appendix 8
Segment plots of characters related to middorsal vertebrae values between Platyceps spp. and related genera (Hemorrhois and Spalerosophis). For abbreviations see Appendix
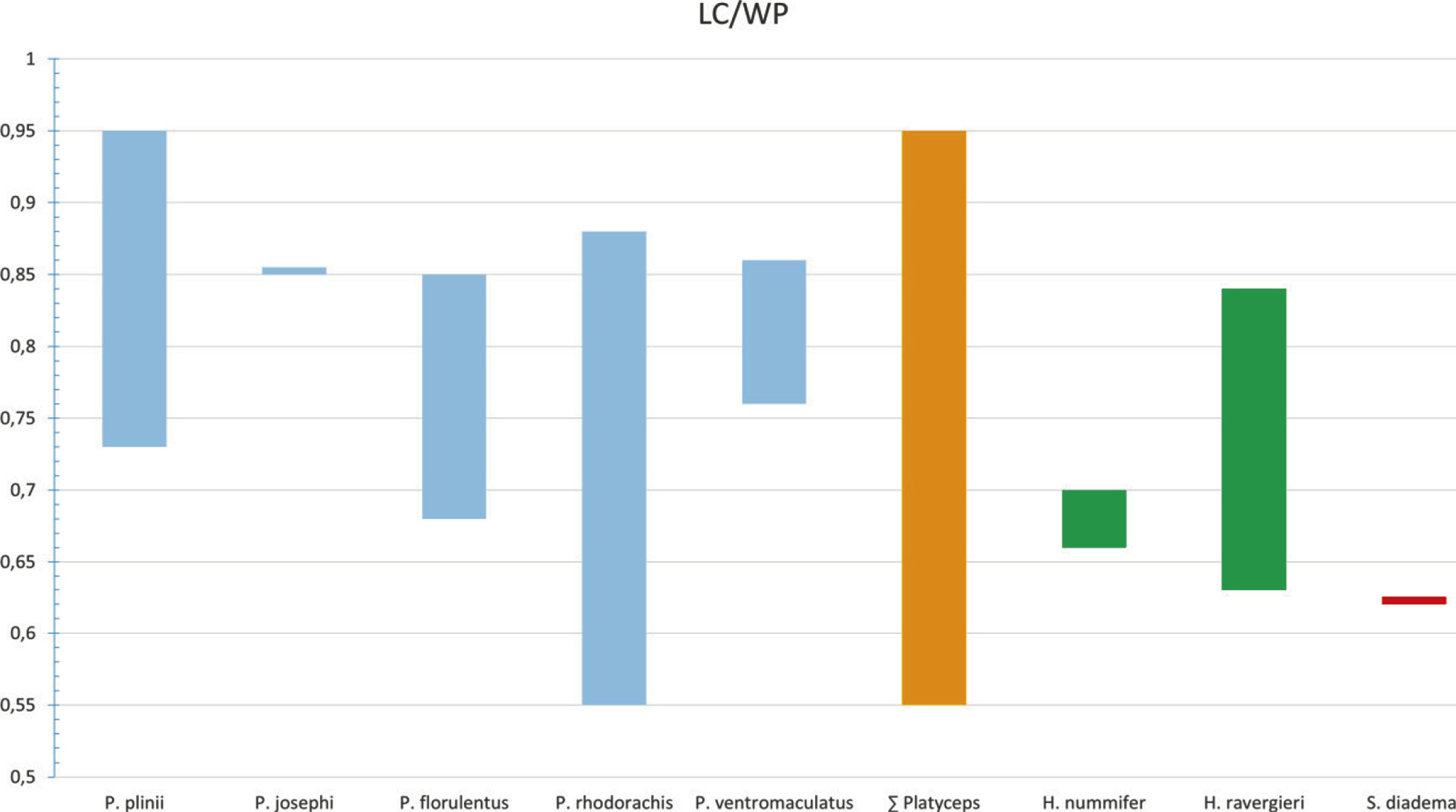
Ratio of the length of centrum to the width across prezygapophyses between outer edges of articular facets (LC/WP).
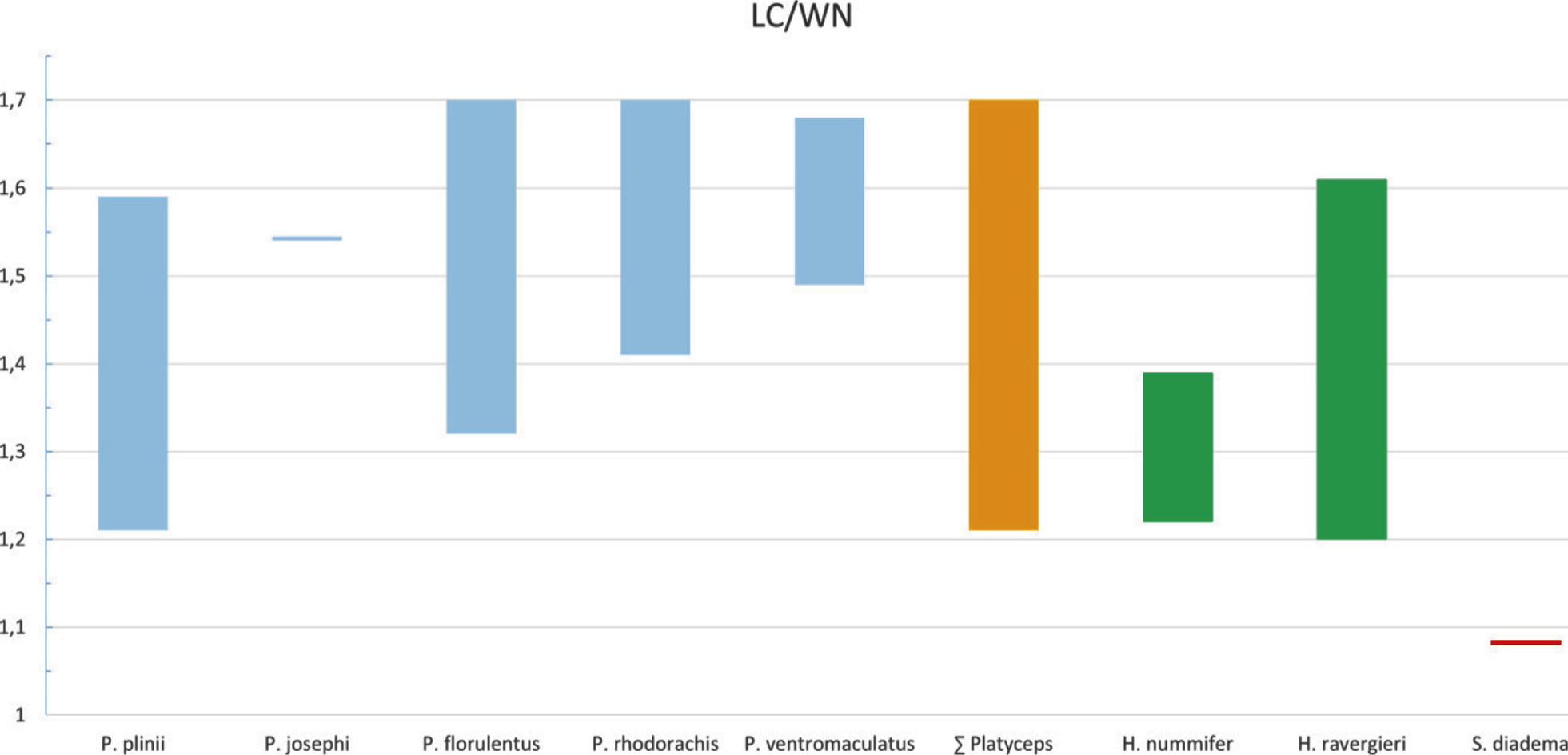
Ratio of the length of centrum and the least width of neural arch (LC/WN).
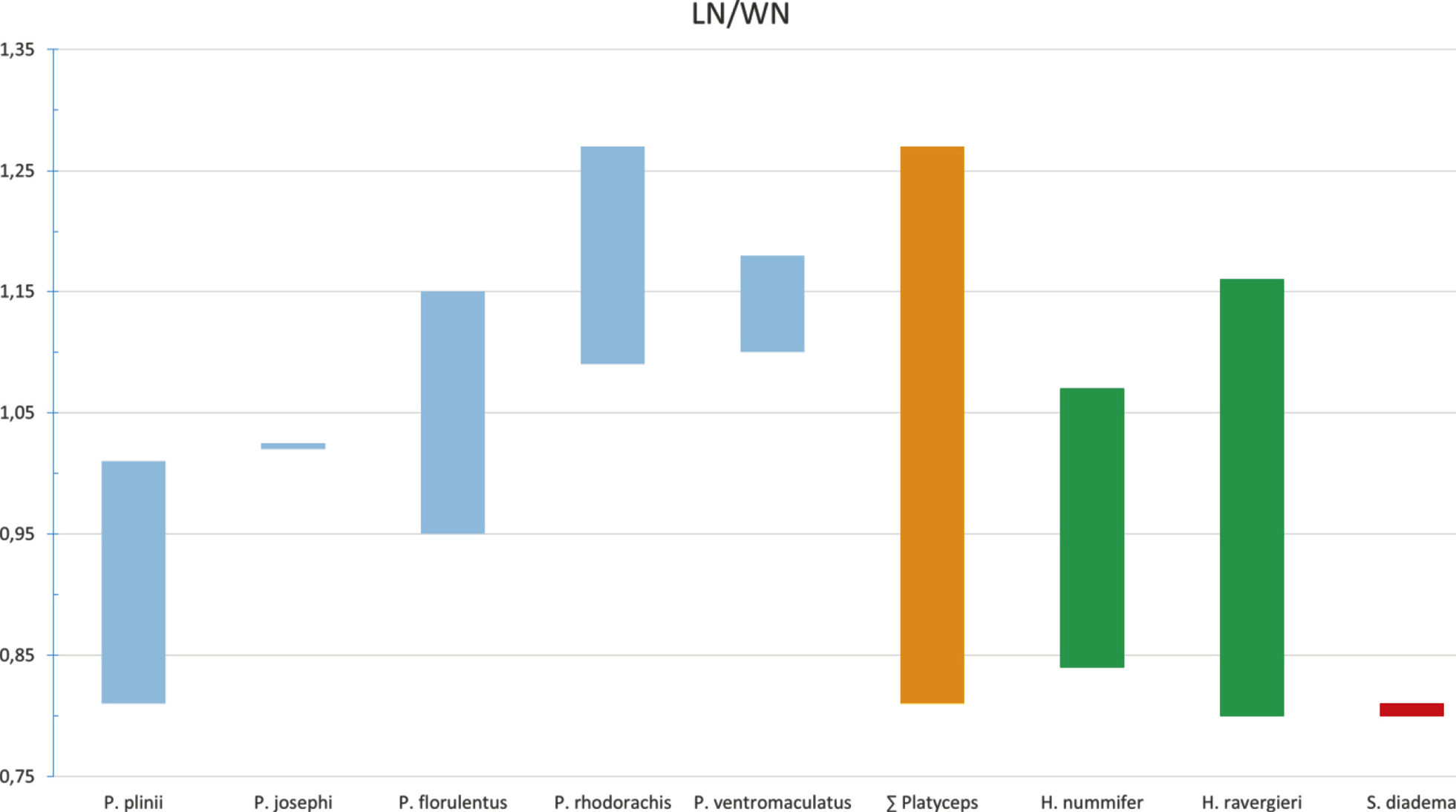
Ratio of length of neural spine to least width of neural arch (LN/WN).
Appendix 9
Gazetteer of confirmed locality records for Platyceps plinii comb. nov. and P. josephi sp. nov. from India and Pakistan mentioned in this study.
| Platyceps plinii comb. nov. | |||||||
|---|---|---|---|---|---|---|---|
| Locality verbatim | Current locality | State | Country | Latitude | Longitude | elevation in m | References |
| Casemcottah [type locality] | Kasimkota | Andhra Pradesh | India | 17.66.660 | 82.967.483 | 26 |
|
| Ellore | Eluru | Andhra Pradesh | India | 16.710425 | 81.115381 | 17 |
|
| Nallamala Hills | Andhra Pradesh | India | 16.283332 | 79.483329 | 350 |
|
|
| Pulicat Lake | Andhra Pradesh | India | 13.640242 | 80.167158 | 1 |
|
|
| Rajamundry North Circars | Rajamundri | Andhra Pradesh | India | 16.983906 | 81.555929 | 67 |
|
| Rollapenta | Rollapenta | Andhra Pradesh | India | 15.889621 | 78.807225 | 474 |
|
| Sundipentha/Sikharam | Sundipenta | Andhra Pradesh | India | 16.076749 | 78.915278 | 470 |
|
| Sunnipenta, Nagarjunasagar Srisailam Tiger Reserve | Sunnipenta | Andhra Pradesh | India | 16.033398 | 78.903697 | 414 |
|
| Thummalapalle uranium mining station, Kadapa and Anantapur districts | Andhra Pradesh | India | 16.947528 | 78.998504 | 250 |
|
|
| Vizagapatam | Visakhapatnam | Andhra Pradesh | India | 17.723127 | 83.301284 | 26 |
|
| Monghyr | Munger | Bihar | India | 25.37556 | 86.47352 | 54 |
|
| Pancht Hill, 2.5 miles from Inanpur, Manbhum | Bihar | India | 23.475107 | 86.048262 | 213 |
|
|
| Patna | Patna | Bihar | India | 25.60932 | 85.12352 | 60 |
|
| Tinpahar, Rajmahal | Rajmahal | Bihar | India | 24.98333 | 87.73333 | 34 |
|
| Kaberdham | Kabirdham | Chhatisgarh | India | 22.095278 | 81.256667 | 365 | This study. |
| Pharsabhar block | Jashpur | Chhatisgarh | India | 22.7702329 | 83.9680735 | 504 |
|
| Bondla Wildlife Sanctuary | Bondla Wildlife Sanctuary | Goa | India | 15.4367676 | 74.1074772 | 199 |
|
| Goa | Goa City | Goa | India | 15.3004543 | 74.0855134 | 90 | Bethencourt Ferreira (1897), |
| Alcock Ashdown & Co., Old Port Bhavnagar | Gujarat | India | 21.7130565 | 71.9608306 | 40 |
|
|
| Baroda | Vadodara | Gujarat | India | 22.3324777 | 73.2174206 | 41 |
|
| Bharuch | Bharuch | Gujarat | India | 21.7518176 | 73.0016822 | 22 |
|
| Bhavnagar | Bhavnagar | Gujarat | India | 21.7669734 | 72.1406835 | 32 |
|
| Dahod City | Dahod | Gujarat | India | 22.8358542 | 74.2556782 | 309 |
|
| Dangs | Dang | Gujarat | India | 20.7413779 | 73.7421809 | 352 |
|
| Gandhinagar | Gandhinagar | Gujarat | India | 23.2248144 | 72.6676020 | 82 |
|
| Gir forest | Gir forest | Gujarat | India | 21.1704377 | 70.7998130 | 278 |
|
| Girnar Hill | Gujarat | India | 21.5178298 | 70.5504226 | 409 |
|
|
| Kala Gadba | Gujarat | India | 21.5270538 | 70.6093521 | 201 |
|
|
| Mehsana | Mehsana | Gujarat | India | 23.5936593 | 72.3893739 | 86 |
|
| near Mahal | Mahal | Gujarat | India | 20.9293702 | 73.6756351 | 318 |
|
| Panchmahals | Panchmahal | Gujarat | India | 22.8205309 | 73.6689945 | 146 |
|
| Sabarkantha | Sabarkantha | Gujarat | India | 24.0294885 | 73.0459884 | 209 |
|
| Samot village, Shoolpaneshwar WS | Samot village, Shoolpaneshwar WS | Gujarat | India | 21.9506182 | 74.3400571 | 199 |
|
| Saurashtra | Gujarat | India | 21.844703 | 70.812761 | 122 | Patel & Vyas (2019). | |
| Surat, suburban residential complex | Gujarat | India | 21.153 | 72.7719 | 8 |
|
|
| Valsad | Valsad | Gujarat | India | 20.5547169 | 72.9961447 | 28 |
|
| Vansda National Park | Gujarat | India | 20.7663784 | 73.4517517 | 144 |
|
|
| Vizapur | Vijapur | Gujarat | India | 23.56015 | 72.7576757 | 126 |
|
| Ambala | Ambala | Haryana | India | 30.3843674 | 76.770421 | 281 |
|
| Hazaribag district | Jharkhand | India | 23.9979667 | 85.3642044 | 608 | Prakash & Raziuddin (2009). | |
| Balekola, Mysore | Bealgola | Karnataka | India | 12.38914 | 76.602571 | 751 | Aengals & Pradhan (2013). |
| Belgaum | Belgaum | Karnataka | India | 15.8572666 | 74.5069343 | 759 |
|
| Bengaluru | Bengaluru | Karnataka | India | 12.9791198 | 77.5912997 | 924 | This study. |
| Chincholi Forest, Kalaburagi district | Karnataka | India | 17.566072 | 77.488044 | 617 |
|
|
| Coorg | Coorg | Karnataka | India | 12.4724402 | 75.7506400 | 951 |
|
| Hosar, near Bangalore | Karnataka | India | 12.715852 | 77.828768 | 873 |
|
|
| Mysore | Mysuru | Karnataka | India | 12.3051828 | 76.6553609 | 752 |
|
| Collagelly Hills, Coimbatore dist | Kollegal Hills | Karnataka | India | 12.1560966 | 77.1071726 | 644 |
|
| Shivamogga City Corporation | Karnataka | India | 13.921822 | 75.566029 | 575 |
|
|
| Muthanga Wildlife Sanctuary | Muthanga, Wayanad district | Kerala | India | 11.69427 | 76.386329 | 868 | Radhakrishnan (1998) |
| Barkatullah University, Bhopal | Madhya Pradesh | India | 23.199352 | 77.451636 | 481 |
|
|
| Bhopal district | Madhya Pradesh | India | 23.265819 | 77.41763 | 497 |
|
|
| Dewas | Dewas | Madhya Pradesh | India | 22.9962075 | 76.1275282 | 540 |
|
| Dumna Nature Park, Jabalpur | Madhya Pradesh | India | 23.169869 | 80.01304 | 462 |
|
|
| Gwalior | Gwalior | Madhya Pradesh | India | 26.2037247 | 78.1573628 | 220 |
|
| Indore | Indore | Madhya Pradesh | India | 22.7203851 | 75.8682103 | 554 |
|
| Jabalpur | Jabalpur | Madhya Pradesh | India | 23.1608938 | 79.9497702 | 420 |
|
| Kanha National Park | Kanha National Park | Madhya Pradesh | India | 22.2766989 | 80.6300919 | 602 | Argawal (1976), |
| Katra Hills, Bhopal | Madhya Pradesh | India | 23.151928 | 77.429774 | 472 |
|
|
| Mandla | Mandla | Madhya Pradesh | India | 22.5986352 | 80.3713545 | 450 |
|
| Pachmarhi Biosphere Reserve | Madhya Pradesh | India | 22.470498 | 78.432348 | 1053 |
|
|
| Satpura Tiger Reserve | Madhya Pradesh | India | 22.502599 | 78.444456 | 968 |
|
|
| Saugor | Sagar | Madhya Pradesh | India | 23.847761 | 78.743331 | 519 |
|
| Shahdol | Shahdol | Madhya Pradesh | India | 23.2804320 | 81.2360533 | 510 |
|
| Sita Hill, Jabalpur | Sita Hill, Jabalpur | Madhya Pradesh | India | 23.1290407 | 79.9433157 | 423 | Sharma, R.C. (1982). |
| Sonemuda (Anuppur District) | Madhya Pradesh | India | 22.662328 | 81.767258 | 972 | Ingle (2020). | |
| Ujjain | Ujjain | Madhya Pradesh | India | 23.174597 | 75.7851423 | 494 |
|
| Ahmednagar | Ahmednagar | Maharashtra | India | 19.1573347 | 74.8602861 | 855 |
|
| Akola district | Maharashtra | India | 20.698656 | 77.094612 | 289 |
|
|
| Ale village, ca. 20 km N of Narayangon | Ale, Pune | Maharashtra | India | 19.1751985 | 74.1154813 | 668 | Sharma, R. C. (1982). |
| Atpadi Tahasil, Sangli district | Maharashtra | India | 17.4130689 | 74.9148231 | 573 |
|
|
| Aurangabad | Aurangabad | Maharashtra | India | 24.7674588 | 84.3719718 | 111 |
|
| Bassein Fort, Thane | Maharashtra | India | 19.329422 | 72.8143531 | 10 |
|
|
| Bhanadara, Nagpur | Bhanadara, Nagpur | Maharashtra | India | 21.1464501 | 80.0483394 | 263 | This study. |
| Bohali | Bohali | Maharashtra | India | 17.6081786 | 75.2511101 | 495 |
|
| Bombay Presidency | Mumbai | Maharashtra | India |
|
|||
| Borivali (Gorai) Mangroves | Maharashtra | India | 19.2393602 | 72.8772926 | 91 |
|
|
| Buldhana district, Amravati division | Maharashtra | India | 20.5739476 | 76.2452802 | 473 |
|
|
| Chink Hill | Chink Hill | Maharashtra | India | 19.5014239 | 75.9872283 | 459 |
|
| Dahanu Forest Division | Maharashtra | India | 19.9887244 | 72.733848 | 15 |
|
|
| Deolali | Deolali, Nasik | Maharashtra | India | 19.9378005 | 73.8564835 | 555 |
|
| Devagad, Sindhudurg district | Maharashtra | India | 16.37512 | 73.38455 | 50 |
|
|
| Ghorawadi, near Talegaon, Pune district | Talegaon, Pune | Maharashtra | India | 18.7395380 | 73.7467945 | 599 |
|
| Jalgaon district | Maharashtra | India | 21.0137606 | 75.5627048 | 218 |
|
|
| Juvem, Salsette | Salsette, Mumbai | Maharashtra | India | 19.167216 | 72.892399 | 171 |
|
| Kaas Plateau | Kas plateau, Satara district | Maharashtra | India | 17.7158767 | 73.8207854 | 1191 |
|
| Katraj Snake Park Pune | Maharashtra | India | 18.4536792 | 73.8563196 | 665 |
|
|
| Khandalla | Khandala | Maharashtra | India | 21.3469486 | 79.3930965 | 297 |
|
| Kolhapur district | Maharashtra | India | 16.6854647 | 74.1368103 | 570 |
|
|
| Kurduvadi | Kurduvadi | Maharashtra | India | 18.090766 | 75.4139381 | 516 |
|
| Maharashtra Nature Park | Mumbai | Maharashtra | India | 19.052278 | 72.862815 | 6 |
|
| Malegaon Tehsil, Washim district | Maharashtra | India | 20.5575459 | 74.5242544 | 432 |
|
|
| Mokhada and Jawhar, Palghar district | Maharashtra | India | 19.9221490 | 73.3311272 | 353 |
|
|
| Nagar | Nagarm, Ahmednagar | Maharashtra | India | 19.1475366 | 74.8027099 | 734 |
|
| Nagpur | Nagpur | Maharashtra | India | 21.1500964 | 79.0127049 | 344 |
|
| Nanded city | Maharashtra | India | 19.0874765 | 77.3279922 | 388 |
|
|
| Navegaon NP, Gondia district | Maharashtra | India | 21.4644517 | 79.1706991 | 331 |
|
|
| Palghar region, Thane | Maharashtra | India | 19.6978751 | 19.6978751 | 47 |
|
|
| Panchgani | Panchgani, Satara | Maharashtra | India | 17.9239543 | 73.7992681 | 1258 |
|
| Parel, Bombay | Parel, Mumbai | Maharashtra | India | 19.0094817 | 72.8376614 | 5 |
|
| Parvati-Pachgaon Hills, Pune | Pune | Maharashtra | India | 18.4969269 | 73.8452906 | 606 |
|
| Poona | Pune | Maharashtra | India | 18.520306 | 73.854318 | 556 |
|
| Powai, Salsette | Powai, Mumbai | Maharashtra | India | 19.1187195 | 72.9073476 | 43 |
|
| Puna | Pune | Maharashtra | India | 18.520306 | 73.854318 | 556 |
|
| Ranidoh region, Pench NP | Pench NP | Maharashtra | India | 21.8854685 | 79.1877054 | 551 |
|
| Santa Cruz, Salsette | Salsette, Mumbai | Maharashtra | India | 19.167216 | 72.892399 | 171 |
|
| Swami Ramanand Teerth Marathwada University, Nanded | Maharashtra | India | 19.100176 | 77.285152 | 384 |
|
|
| Thana | Thane | Maharashtra | India | 19.194329 | 72.970177 | 20 |
|
| Uran wetland, Navi, Mumbai | Mumbai | Maharashtra | India | 19.07283 | 72.88261 | 8 |
|
| Vidarbha region | Maharashtra | India | 21.0163970 | 78.9863482 | 291 |
|
|
| Vidyanagari campus Mumbai Uni, Maharashtra | Vidyanagari Campus | Maharashtra | India | 20.9702409 | 74.8026808 | 258 |
|
| Visapur | Visapur, Ahmednagar | Maharashtra | India | 20.9702409 | 73.4927555 | 115 |
|
| Yavatmal | Yavatmal | Maharashtra | India | 20.3367414 | 78.2295788 | 377 |
|
| Marol, Mumbai | Mumbai | Mahrashtra | India | 19.116801 | 72.884675 | 15 |
|
| Baripada | Baripada | Odisha | India | 21.9207475 | 86.8553154 | 64 | This study. |
| Choudwar, Cuttack | Choudwar, Cuttack | Odisha | India | 20.5276823 | 85.910155 | 32 | This study. |
| Gandhamardan Hills | Odisha | India | 20.8609068 | 82.7775689 | 293 |
|
|
| Nandankanan Biological Park | Nandankanan Biological Park. Cuttack | Odisha | India | 20.3977336 | 85.8138633 | 36 |
|
| Patharakali area, Bhadrak district | Odisha | India | 20.973825 | 86.615244 | 19 |
|
|
| Pondicherry | Pondicherry | Puducherry | India | 11.820894 | 79.697454 | 14 | This study. |
| near Hemavas Dam, Pali district | Hemwas, Pali | Rajasthan | India | 25.7456159 | 73.3473000 | 220 |
|
| Pratapgarh district | Rajasthan | India | 23.9936599 | 74.7284642 | 459 | Anonymous (n. d., p. 13) | |
| Ramri | Rajasthan | India | 26.04698 | 76.297146 | 253 |
|
|
| Farahabad | Telangana state | India | 16.282541 | 78.680858 | 537 |
|
|
| Hyderabad Region | Telangana state | India | 17.344307 | 78.531531 | 486 | Walmiki) 2012b), |
|
| Ippalapally | Telangana state | India | 16.354054 | 79.074511 | 419 |
|
|
| Jannaram | Telangana state | India | 19.115557 | 78.999515 | 190 |
|
|
| Kerimeri | Telangana state | India | 19.44283 | 79.051793 | 277 |
|
|
| Khanapur | Telangana state | India | 19.041383 | 78.648385 | 248 |
|
|
| Kinnerasani | Telangana state | India | 17.67149 | 80.676419 | 94 |
|
|
| Old Bowenpally | Telangana state | India | 17.481334 | 78.478662 | 548 |
|
|
| OU Tarnaka | Telangana state | India | 17.415796 | 78.533936 | 517 |
|
|
| Rushulacheruvu | Telangana state | India | 14.421364 | 77.801929 | 354 |
|
|
| Vijayapuri | Telangana state | India | 16.589441 | 79.335993 | 149 |
|
|
| Allahabad | Prayagraj | Uttar Pradesh | India | 25.4381302 | 81.8338005 | 103 |
|
| Benares | Varanasi | Uttar Pradesh | India | 25.3356491 | 83.0076292 | 79 |
|
| Fyzabad | Faizabad | Uttar Pradesh | India | 26.7431323 | 82.0898131 | 105 |
|
| Mathura region | Uttar Pradesh | India | 27.491322 | 77.688805 | 165 |
|
|
| Calcutta | Kolkata | West Bengal | India | 22.5677459 | 88.3476023 | 10 |
|
| Dum Dum | West Bengal | India | 22.647.051 | 88.431.686 | 10 |
|
|
| Durgapur city | Durgapur | West Bengal | India | 23,5788078 | 87,3264641 | 88 |
|
| Hosur Hills | Tamil Nadu | India | 12.739657 | 77.856756 | 851 |
|
|
| Tambaram, Chennai | Tamil Nadu | India | 12.918878 | 80.119084 | 36 |
|
|
| Tambaram, Chennai | Tamil Nadu | India | 12.918878 | 80.119084 | 36 |
|
|
| Lal Suhanra National Park, Bahawalpur | Bahawalpur | Punjab | Pakistan | 29.313483 | 71.905294 | 125 |
|
| Badin | Badin | Sindh | Pakistan | 24.6332316 | 68.88634238 | 10 |
|
| Hala | Hala | Sindh | Pakistan | 25.5971858 | 68.4454874 | 25 |
|
| Jati | Jati | Sindh | Pakistan | 24.3533505 | 68.2678116 | -50 |
|
| Jogis | Jogi | Sindh | Pakistan | 27.67621 | 68.216521 | 50 |
|
| Larkana district | Larkana | Sindh | Pakistan | 27.5564798 | 68.2101509 | 48 |
|
| Makli Hills near Tatta | Makli hills nr. Tatta | Sindh | Pakistan | 24.7529974 | 67.9002813 | 30 |
|
| Mohenjo-daro | Mohenja-daro | Sindh | Pakistan | 27.3268899 | 68.1371029 | 52 |
|
| Pir Patho | Pir Patho, Thatta | Sindh | Pakistan | 24.55964 | 67.88535 | 20 |
|
| Raj Malk, Thatta district, Mirpur Sakro Area | Raj Malk, Thatta | Sindh | Pakistan | 24.7459022 | 67.9465502 | 7 |
|
| Sonda (1 mile east of) | Sonda, Thatta district | Sindh | Pakistan | 24.9865058 | 68.7537357 | 15 |
|
| Tatta (4 miles east of) | Thatta | Sindh | Pakistan | 24.7486460 | 67.8235033 | 10 |
|
| Platyceps josephi sp. nov. | |||||||
| Anaikatty, Coimbatore | Anaikatti, Coimbatore | Tamil Nadu | India | 11.114903 | 76.756551 | 582 | This study. |
| Anamalai Hills | Anamalai hills | Tamil Nadu | India | 10.479552 | 77.17458 | 404 |
|
| Auroville, Villupuram | Auroville, Villupuram | Tamil Nadu | India | 12.005421 | 79.8111 | 52 |
|
| Batlagundu, Dindugal district | Batlagundu, Dindugal | Tamil Nadu | India | 10.163609 | 77.759098 | 237 | This study. |
| Coimbatore | Coimbatore `by implication’ | Tamil Nadu | India | 10.973923 | 77.110244 | 415 |
|
| Coutrallam | Courtallam | Tamil Nadu | India | 8.924261 | 77.289739 | 190 | This study. |
| Karur | Karur | Tamil Nadu | India | 10.960405 | 78.077561 | 133 | This study. |
| Madurai | Madurai | Tamil Nadu | India | 9.925201 | 78.119775 | 139 |
|
| Manimutharu | Tirunelveli | Tamil Nadu | India | 8.654005 | 77.425398 | 79 | This study. |
| Maruthuvazhmalai, Kanyakumari | Maruthuvazhmalai, Kanyakumari | Tamil Nadu | India | 8.124244 | 77.512661 | 243 |
|
| Meghamalai Hills | Meghamalai Hills | Tamil Nadu | India | 9.6678528 | 77.283688 | 445 |
|
| Pollachi I | Pollachi | Tamil Nadu | India | 10.658823 | 77.00873 | 293 | This study. |
| Pollachi II | Pollachi | Tamil Nadu | India | 10.746908 | 77.089955 | 342 | This study. |
| Salem | Salem | Tamil Nadu | India | 11.741785 | 77.938889 | 314 | This study. |
| Tiruneveli | Tirunelveli | Tamil Nadu | India | 8.7012212 | 77.776077 | 44 |
|
| Trichinopoly | Tiruchirapalli | Tamil Nadu | India | 10.804973 | 78.686904 | 84 |
|
| Tuticorin (type locality) | Thoothukudi | Tamil Nadu | India | 8.8052602 | 78.145275 | 7 |
|
| Vadipatti, Madurai dist | Vadipatti, Madurai dist | Tamil Nadu | India | 10.11222 | 77.97659 | 214 | This study. |
Appendix 10
Material examined
(Material examined for this study. Symbol * denotes for specimens CT scanned for analysis of the osteology and symbol ° for specimens used in analysis of dorsal scale reduction.)
Hemorrhois ravergieri (1)
Without specified locality:
Lycodon cf. aulicus (7)
INDIA – without specified locality, Russell’s dried snake skin collection:
Lycodon fasciolatus (15)
INDIA – Andhra Pradesh: ZSI-CZRC-V 6902 from Bellupada, Ichchapuram; Odisha: ZSI-CZRC-V 6411, 6447, 6450, 6451, 6460, 6463, 6464 and 6530, all from Ganjam; without specified locality:
Platyceps bholanathi (3)
ZSI-K 21337, Nagarjuna hills, Andhra Pradesh (Holotype); ZSI-K 21335–36, Nagarjuna hills, Andhra Pradesh (paratypes)
Platyceps florulentus (1)
EGYPT –
Platyceps josephi sp. nov. (11)
INDIA –Tamil Nadu:
Platyceps plinii comb. nov. (n=50)
PAKISTAN – Sindh:
INDIA – Andhra Pradesh:
Platyceps rhodorachis (4)
TURKMENISTAN –
Platyceps ventromaculatus (1)
PAKISTAN – Sindh:
Spalerosophis diadema (1)
Without specified locality: